وµ·ه؛•ه †ç©چ物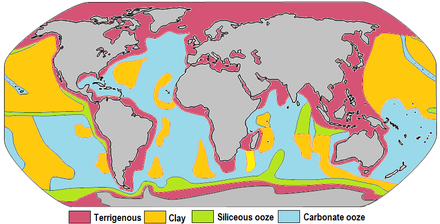 وµ·ه؛•ه †ç©چ物(مپ‹مپ„مپ¦مپ„مپںمپ„مپ›مپچمپ¶مپ¤ï¼‰م€پمپ¾مپںمپ¯وµ·و´‹ه †ç©چ物(مپ‹مپ„م‚ˆمپ†مپںمپ„مپ›مپچمپ¶مپ¤ï¼‰مپ¯م€پوµ·ه؛•مپ«è“„ç©چمپ—مپںن¸چو؛¶و€§ç²’هگمپ®ه †ç©چ物مپ®مپ“مپ¨مپ§مپ‚م‚‹م€‚مپ“م‚Œم‚‰مپ®ç²’هگمپ¯م€پهœںه£Œم‚„ه²©çں³مپ«ç”±و¥مپ—陸هœ°مپ‹م‚‰ن¸»مپ«و²³ه·م‚’経由مپ—مپ¦وµ·مپ¾مپ§éپ‹وگ¬مپ•م‚Œمپ¦مپچمپںم‚‚مپ®م‚„م€پ風مپ«م‚ˆمپ£مپ¦éپ‹مپ°م‚Œمپںه،µم€پو°·و²³مپھمپ©مپ«م‚ˆمپ£مپ¦éپ‹مپ°م‚Œوµ·مپ«وµپم‚Œè¾¼م‚€م‚‚مپ®مپŒهگ«مپ¾م‚Œم‚‹م€‚مپ¾مپںم€پوµ·و´‹ç”ں物م‚„وµ·و°´ن¸مپ®هŒ–ه¦çڑ„و²ˆو®؟物م€پمپٹم‚ˆمپ³وµ·ه؛•çپ«ه±±م‚„éڑ•çں³مپ®و®‹éھ¸مپھمپ©م‚‚هگ«مپ¾م‚Œمپ¦مپ„م‚‹م€‚ ن¸ه¤®وµ·ه¶؛مپ®و•°kmè؟‘è¾؛مپھمپ©مپ®çپ«ه±±ه²©مپŒه¤ڑمپڈ見م‚‰م‚Œم‚‹ن¾‹ه¤–çڑ„مپھه ´و‰€م‚’除مپ„مپ¦م€پوµ·ه؛•مپ®ه¤§هچٹمپ¯ه †ç©چ物مپ«م‚ˆمپ£مپ¦è¦†م‚ڈم‚Œمپ¦مپ„م‚‹م€‚ه †ç©چ物م‚’و§‹وˆگمپ™م‚‹وˆگهˆ†مپ®ç”±و¥و؛گمپ¯è¤‡و•°مپ‚م‚ٹم€پمپمپ®çµ„وˆگمپ¯éه¸¸مپ«ه¤ڑو§کمپ§مپ‚م‚‹م€‚وµ·ه؛•مپ®ه †ç©چ物مپ®هژڑمپ•مپ¯م€پو•°mmمپ‹م‚‰و•°هچپkmمپ«هڈٹمپ¶م€‚وµ·ه؛•ه †ç©چ物مپ¯وµ·é¢è؟‘مپڈمپ§مپ¯وœھه›؛çµگمپ®مپ¾مپ¾مپ§مپ‚م‚‹مپŒم€پو•°ç™¾mمپ‹م‚‰و•°هچƒmمپ®و·±مپ•مپ§مپ¯و°´هœ§مپ®مپںم‚په †ç©چ物مپŒçں³هŒ–(ه²©çں³هŒ–)مپ™م‚‹م€‚ ه †ç©چ物مپ®è“„ç©چé€ںه؛¦مپ¯م€پوµ·و´‹مپ®ه¤§éƒ¨هˆ†مپ§مپ¯éه¸¸مپ«éپ…مپڈم€پوکژç¢؛مپھه †ç©چ物مپ®ه±¤مپŒه½¢وˆگمپ•م‚Œم‚‹مپ¾مپ§مپ«و•°هچƒه¹´م‚‚مپ®و™‚é–“مپŒمپ‹مپ‹م‚‹م€‚陸هœ°مپ‹م‚‰éپ‹مپ°م‚Œمپںه †ç©چ物مپ¯وœ€م‚‚é€ںمپڈè“„ç©چمپ—م€پç²—مپ„ç²’هگمپ®ه ´هگˆمپ¯1000ه¹´مپ‚مپںم‚ٹ1mن»¥ن¸ٹمپ®é€ںه؛¦مپ§ه †ç©چمپ™م‚‹مپ“مپ¨م‚‚مپ‚م‚‹م€‚مپ¾مپںم€پوµپé‡ڈمپ®ه¤ڑمپ„ه¤§و²³ه·مپ®و²³هڈ£ن»کè؟‘مپھمپ©مپ§مپ¯م€په †ç©چé€ںه؛¦مپŒمپ•م‚‰مپ«و،پéپ•مپ„مپ«و—©مپڈمپھم‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹م€‚ن¸€و–¹مپ§م€پç”ں物起و؛گمپ®è»ںو³¥مپھمپ©مپ¯ 1000ه¹´مپ«ç´„1cm程ه؛¦مپ®é€ںه؛¦مپ§è“„ç©چمپ—م€په°ڈمپ•مپھç²کهœںç²’هگمپ«è‡³مپ£مپ¦مپ¯1000ه¹´مپ«ç´„1mm程ه؛¦مپ®é€ںه؛¦مپ§ه †ç©چمپ™م‚‹م€‚ 陸هœ°مپ‹م‚‰مپ®ه †ç©چ物مپ¯م€پهœ°è،¨وµپه‡؛م€پو²³ه·وµپه‡؛م€پمپٹم‚ˆمپ³مپمپ®ن»–مپ®مƒ—مƒم‚»م‚¹مپ«م‚ˆمپ£مپ¦ه¤§é™¸و–œé¢مپ«ه †ç©چمپ—م€پمپ•م‚‰مپ«و؟پوµپمپ«م‚ˆمپ£مپ¦ه¤§é™¸و–œé¢مپ‹م‚‰و·±وµ·ه؛•مپ¾مپ§éپ‹مپ°م‚Œم‚‹م€‚مپمپ—مپ¦و·±وµ·ه؛•مپ«è“„ç©چمپ•م‚Œمپںه †ç©چ物مپ¯م€پن¸ه¤®وµ·ه¶؛مپ‹م‚‰ه؛ƒمپŒم‚‹مƒ—مƒ¬مƒ¼مƒˆمپ®ه‹•مپچمپ«ه¾“مپ£مپ¦م€پهœ°çگƒمپ®ه†…部مپ«م‚†مپ£مپڈم‚ٹمپ¨و²ˆمپ؟è¾¼م‚“مپ§مپ„مپڈم€‚ه†…部مپ«هڈ–م‚ٹè¾¼مپ¾م‚Œمپںه †ç©چ物مپ¯و؛¶èچمپ—م€پمپمپ®ن¸€éƒ¨مپ¯و؛¶ه²©وµپم‚„熱و°´ه™´ه‡؛ه”مپ‹م‚‰و”¾ه‡؛مپ•م‚Œم‚‹مپ“مپ¨مپ§ه†چمپ³هœ°è،¨مپ¸مپ¨وˆ»م‚‹م€‚ ه †ç©چ物مپ¯م€په¤ڑو•°مپ®ه؛•ç”ںوµ·و´‹ç”ں物م€پ特مپ«وµ·و´‹ه¾®ç”ں物مپ«مپ¨مپ£مپ¦é‡چè¦پمپھç”ںوپ¯ه ´و‰€مپ¨مپھمپ£مپ¦مپ„م‚‹م€‚ه †ç©چ物مپ®هŒ–çں³مپ«مپ¯م€پéپژهژ»مپ®و°—ه€™م€پمƒ—مƒ¬مƒ¼مƒˆمƒ†م‚¯مƒˆمƒ‹م‚¯م‚¹م€پوµ·و´‹ه¾ھç’°مƒ‘م‚؟مƒ¼مƒ³م€پن¸»è¦پمپھ絶و»…مپ®م‚؟م‚¤مƒںمƒ³م‚°مپھمپ©مپ«é–¢مپ™م‚‹وƒ…ه ±مپŒهگ«مپ¾م‚Œمپ¦مپٹم‚ٹم€په¦è،“çڑ„مپ«م‚‚é‡چè¦پمپھç ”ç©¶é،Œوگمپ§مپ‚م‚‹[1]م€‚ و¦‚è¦پو¯”較çڑ„è‹¥مپ„çپ«ه±±ه²©مپ«م‚ˆمپ£مپ¦و§‹وˆگمپ•م‚Œم‚‹ن¸ه¤®وµ·ه¶؛مپ®و•°kmè؟‘è¾؛مپ¨مپ„مپ£مپںè‹¥ه¹²مپ®ن¾‹ه¤–م‚’除مپ„مپ¦م€پوµ·ه؛•مپ®مپ»مپ¨م‚“مپ©مپ®éƒ¨هˆ†مپ¯ه †ç©چ物مپ§è¦†م‚ڈم‚Œمپ¦مپ„م‚‹م€‚ه †ç©چ物م‚’و§‹وˆگمپ™م‚‹ç‰©è³ھمپ®ç”±و¥مپ¯و§کم€…مپ§مپ‚م‚ٹم€په¤§é™¸مپ¸مپ®è؟‘مپ•م‚„و°´و·±م€پوµ·وµپم€پç”ں物و´»ه‹•م€پو°—ه€™مپھمپ©مپ«ه؟œمپکمپ¦çµ„وˆگمپŒه¤§مپچمپڈç•°مپھم‚‹م€‚وµ·ه؛•ه †ç©چ物 (مپٹم‚ˆمپ³ه †ç©چه²©) مپ®هژڑمپ•مپ¯م€پو•°mmمپ‹م‚‰و•°هچپkmمپ«هڈٹمپ¶م€‚è،¨é¢è؟‘مپڈمپ§مپ¯م€پوµ·ه؛•ه †ç©چ物مپ¯وœھه›؛çµگمپ®مپ¾مپ¾مپ§مپ‚م‚‹مپŒم€په †ç©چ物مپ®ç¨®é،م‚„مپمپ®ن»–مپ®è¦په› مپ«م‚ˆمپ£مپ¦ç•°مپھم‚‹م‚‚مپ®مپ®م€پو•°ç™¾مپ‹م‚‰و•°هچƒmمپ®و·±مپ•مپ§ ه †ç©چ物مپ¯çں³هŒ–مپ™م‚‹[2]م€‚وµ·ه؛•ه †ç©چ物مپ®ç”±و¥مپ¯و¬،مپ®م‚ˆمپ†مپ«è¦پç´„مپ§مپچم‚‹[2]م€‚
مپ“م‚Œم‚‰مپ®ç‰©è³ھمپ®وµ·هںںهˆ†ه¸ƒمپ¯م€پمپ“مپ®è¨کن؛‹مپ®ه†’é مپ®ه›³مپ«ç¤؛مپ—مپںمپ¨مپٹم‚ٹمپ§مپ‚م‚‹م€‚陸ç”ںه †ç©چ物مپ¯م€په¤§é™¸مپ®è؟‘مپڈم‚„ه†…وµ·م€په¤§مپچمپھو¹–مپ®ن¸مپھمپ©مپ§ه„ھه‹¢مپ§مپ‚م‚‹م€‚مپ“م‚Œم‚‰مپ®ه †ç©چ物م‚’و§‹وˆگمپ™م‚‹ç²’هگمپ¯و¯”較çڑ„ç²—مپ„ه‚¾هگ‘مپŒمپ‚م‚ٹم€پé€ڑه¸¸مپ¯ç ‚مپ¨م‚·مƒ«مƒˆم‚’هگ«م‚€مپŒم€په ´هگˆمپ«م‚ˆمپ£مپ¦مپ¯ه°ڈçں³م‚„çژ‰çں³مپŒهگ«مپ¾م‚Œم‚‹م€‚ç²کهœںمپ¯و²؟ه²¸ç’°ه¢ƒمپ«ه®ڑç€مپ™م‚‹م‚‚مپ®م‚‚مپ‚م‚‹مپŒم€په¤ڑمپڈمپ¯وµ·وµپمپ«م‚ˆمپ£مپ¦مپمپ®ç™؛ç”ںو؛گمپ‹م‚‰éپ مپڈ離م‚Œمپںه ´و‰€مپ¸مپ¨هˆ†و•£مپ™م‚‹م€‚ç²کهœں鉱物مپ¯م€پو·±وµ·مپ®ه؛ƒمپ„هœ°هںںمپ§ه„ھه‹¢مپ§مپ‚م‚ٹم€پمپ“مپ®ç²کهœںمپ®مپ»مپ¨م‚“مپ©مپ¯ه¤§é™¸èµ·و؛گمپ§مپ‚م‚‹م€‚çڈھè³ھè»ںو³¥ (ن¸»مپ«و”¾و•£è™«مپ¨çڈھè—»مپ«ç”±و¥) مپ¯م€پهچ—و¥µهœ°هںںم‚„ه¤ھه¹³و´‹مپ®èµ¤éپ“و²؟مپ„م€پم‚¢مƒھمƒ¥مƒ¼م‚·مƒ£مƒ³هˆ—ه³¶مپ®هچ—م€پم‚¤مƒ³مƒ‰و´‹مپ®ه¤§éƒ¨هˆ†مپھمپ©مپ§م‚ˆمپڈ見م‚‰م‚Œم‚‹م€‚ç‚é…¸ه،©مپ®و»²ه‡؛物مپ¯م€پ赤éپ“مپٹم‚ˆمپ³ن¸ç·¯ه؛¦هœ°هںںه†…مپ®مپ™مپ¹مپ¦مپ®وµ·و´‹مپ«ه؛ƒمپڈهˆ†ه¸ƒمپ—مپ¦مپ„م‚‹م€‚ه®ںéڑ›م€پç²کهœںمپ¯وµ·مپ®مپ„مپںم‚‹ه ´و‰€مپ®ه †ç©چ物مپ§è¦³ه¯ںمپ•م‚Œم‚‹مپŒم€پم‚·مƒھم‚«مپ¨ç‚é…¸ه،©م‚’ç”ںوˆگمپ™م‚‹ç”ں物مپŒè±ٹه¯Œمپھوµ·هںںمپ§مپ¯م€پç²کهœںن»¥ن¸ٹمپ«ه¤ڑمپڈمپ®م‚·مƒھم‚«م‚„ç‚é…¸ه،©م‚’هگ«م‚€ه †ç©چ物م‚’ç”ںوˆگمپ™م‚‹[2]م€‚ ç‚é…¸ه،©ه †ç©چ物مپ¯م€پç‚é…¸ه،©مپ‹م‚‰و®»م‚’ن½œم‚‹م€پوµ·و´‹è،¨ه±¤مپ§ه؛ƒç¯„ه›²مپ«هکهœ¨مپ™م‚‹éپ و´‹ç”ں物مپ«ç”±و¥مپ™م‚‹م€‚مپ“م‚Œم‚‰مپ®ه°ڈمپ•مپھو®»م‚„م€پç •مپ‘مپںمپ¨مپچمپ«ه½¢وˆگمپ•م‚Œم‚‹مپ•م‚‰مپ«ه°ڈمپ•مپھç ´ç‰‡مپ¯م€پو°´وں±م‚’é€ڑمپ£مپ¦م‚†مپ£مپڈم‚ٹمپ¨و²ˆé™چمپ™م‚‹مپŒم€په؟…مپڑمپ—م‚‚ه؛•مپ«هˆ°éپ”مپ™م‚‹مپ¨مپ¯é™گم‚‰مپھمپ„م€‚مپ“م‚Œمپ¯م€پو–¹è§£çں³مپ¯هœ°è،¨مپ®و°´مپ«مپ¯و؛¶è§£مپ—مپھمپ„مپŒم€پمپمپ®و؛¶è§£ه؛¦مپ¯و·±مپ• (هœ§هٹ›) مپ¨مپ¨م‚‚مپ«ه¢—هٹ مپ™م‚‹مپںم‚پم€پ4,000mن»کè؟‘مپ§مپ¯ç‚é…¸ه،©مپ®ç ´ç‰‡مپ¯وµ·و°´ن¸مپ¸مپ¨و؛¶è§£مپ™م‚‹مپںم‚پمپ§مپ‚م‚‹م€‚مپ“مپ®و°´و·±مپ¯ç‚é…¸ه،©è£œه„ںو·±ه؛¦ï¼ˆCCD)مپ¨مپ—مپ¦çں¥م‚‰م‚Œمپ¦مپٹم‚ٹم€پç·¯ه؛¦مپ¨و°´و¸©مپ«م‚ˆمپ£مپ¦ه¤‰هŒ–مپ™م‚‹م€‚مپمپ®çµگوœم€پوµ·مپ®وœ€م‚‚و·±مپ„部هˆ† (4,000mن»¥و·±) مپ«مپ¯ç‚é…¸ه،©مپ®و»²ه‡؛مپ¯è¦‹م‚‰م‚Œمپھمپ„مپŒم€په¤§è¥؟و´‹ن¸ه¤®وµ·ه¶؛م‚„و±ه¤ھه¹³و´‹وµ·è†¨ (هچ—م‚¢مƒ،مƒھم‚«مپ®è¥؟) م€پمƒڈمƒ¯م‚¤ه¤©çڑ‡وµ·ه±±ç¾¤ (هŒ—ه¤ھه¹³و´‹)م€پمپٹم‚ˆمپ³ه¤ڑمپڈمپ®ه¤ç«‹مپ—مپںوµ·ه±±مپ®é ‚ن¸ٹمپھمپ©مپ®وµ…وµ·هںںمپ§مپ¯م‚ˆمپڈ見م‚‰م‚Œم‚‹[2]م€‚ ه †ç©چ物組織 ç²’ه؛¦éڑژهŒ؛هˆ†ï¼ˆم‚¦م‚§م‚¹مƒˆمƒ³مƒ¯مƒ¼م‚¹م‚¹م‚±مƒ¼مƒ«ï¼‰مپ«هں؛مپ¥مپ„مپ¦ç²’هگم‚µم‚¤م‚؛مپ«م‚ˆم‚ٹهˆ†é،مپ•م‚Œمپںه †ç©چ物هŒ؛هˆ† ه †ç©چ物مپ®çµ„織(مƒ†م‚¯م‚¹مƒپمƒ£ï¼‰مپ®è©•ن¾،و–¹و³•مپ«مپ¯مپ„مپڈمپ¤مپ‹مپ®ç¨®é،مپŒمپ‚م‚‹م€‚ ن¸€مپ¤ç›®مپ¯ç²’ه؛¦مپ§مپ‚م‚ٹم€پç²’ه؛¦éڑژهŒ؛هˆ†ï¼ˆم‚¦م‚§مƒ³مƒˆمƒ¯مƒ¼م‚¹م‚¹م‚±مƒ¼مƒ«ï¼‰مپ«ه¾“مپ£مپ¦ç²’هگم‚µم‚¤م‚؛مپ§هˆ†é،مپ™م‚‹مپ“مپ¨مپŒمپ§مپچم‚‹[1]م€‚ن¾‹مپˆمپ°م€پç²کهœںمپ¯ç²’ه¾„مپŒ 0.004mmوœھو؛€مپ®وœ€م‚‚ç´°مپ‹مپ„م‚‚مپ®م‚’وŒ‡مپ—م€په·¨ç¤«مپ¯وœ€ه¤§مپ§ç²’ه¾„مپ¯ 256mmن»¥ن¸ٹمپ®م‚‚مپ®م‚’وŒ‡مپ™[3]م€‚ç²’هگم‚µم‚¤م‚؛مپ¯ن»–مپ®ه› هگمپ¨و¯”較مپ—مپ¦م€په †ç©چ物مپŒه †ç©چمپ™م‚‹و،ن»¶م‚’وœ€م‚‚م‚ˆمپڈèھ¬وکژمپ§مپچم‚‹م€‚ن¾‹مپˆمپ°ه¼·مپ„وµپم‚Œم‚„و³¢مپھمپ©مپ®é«کم‚¨مƒچمƒ«م‚®مƒ¼مپŒهکهœ¨مپ™م‚‹و،ن»¶ن¸‹مپ§مپ¯é€ڑه¸¸م€پç´°مپ‹مپ„ç²’هگمپŒéپ‹مپ³هژ»م‚‰م‚Œمپ¦مپ—مپ¾مپ†مپںم‚پم€په¤§مپچمپھç²’هگمپ®مپ؟مپŒه †ç©چمپ™م‚‹م€‚逆مپ«م‚ˆم‚ٹن½ژمپ„م‚¨مƒچمƒ«م‚®مƒ¼و،ن»¶ن¸‹مپ§مپ¯م€پم‚ˆم‚ٹه°ڈمپ•مپھç²’هگمپŒو²ˆé™چمپ—م€پم‚ˆم‚ٹç´°مپ‹مپ„ه †ç©چ物م‚’ه½¢وˆگمپ™م‚‹[1]م€‚   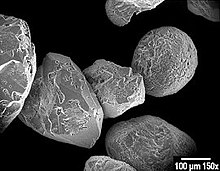 2مپ¤ç›®مپ¯ç²’هگم‚µم‚¤م‚؛مپ®هˆ†ه¸ƒï¼ˆم‚½مƒ¼مƒ†م‚£مƒ³م‚°م€پSorting)مپ§مپ‚م‚ٹم€په †ç©چ物مپ®مƒ†م‚¯م‚¹مƒپمƒ£م‚’هˆ†é،مپ—ç²’هگم‚µم‚¤م‚؛مپ®ه‡ن¸€و€§مپ‹م‚‰è©•ن¾،مپ•م‚Œم‚‹م€‚ç ‚وµœمپ®ç ‚مپ®م‚ˆمپ†مپ«مپ™مپ¹مپ¦مپ®ç²’هگمپŒهگŒو§کمپ®م‚µم‚¤م‚؛مپ§مپ‚م‚‹ه ´هگˆم€په †ç©چ物مپ¯م‚ˆمپڈéپ¸هˆ¥مپ•م‚Œمپ¦مپ„م‚‹مپ¨è¨€مپˆم‚‹مپŒم€پ逆مپ«ن¾‹مپˆمپ°و°·و²³ه †ç©چ物مپ®م‚ˆمپ†مپ«ç²’هگم‚µم‚¤م‚؛مپŒو§کم€…مپ§مپ‚م‚‹ه ´هگˆم€په †ç©چ物مپ®éپ¸هˆ¥مپŒن¸چهچپهˆ†مپ§مپ‚م‚‹مپ¨è¨€مپˆم‚‹[1]م€‚ 3 مپ¤ç›®مپ®و–¹و³•مپ¯م€پمپمپ®وˆگç†ںه؛¦م€پمپ™مپھم‚ڈمپ،مپمپ®ç²’هگمپŒو°´مپ«م‚ˆمپ£مپ¦éپ‹مپ°م‚Œمپںو™‚é–“مپ®ه½±éں؟م‚’èھ؟مپ¹م‚‹مپ“مپ¨مپ§مپ‚م‚‹م€‚مپ“مپ®وˆگç†ںه؛¦م‚’ç¤؛مپ™ 1 مپ¤مپ®و–¹و³•مپ¯م€پç²’هگمپ®ن¸¸مپ•م‚’èھ؟مپ¹م‚‹مپ“مپ¨مپ§مپ‚م‚‹م€‚ه †ç©چ物مپŒوˆگç†ںمپ™م‚Œمپ°مپ™م‚‹مپ»مپ©م€پو™‚é–“مپ®çµŒéپژمپ¨مپ¨م‚‚مپ«و‘©è€—مپ—م€پمپمپ®çµگوœç²’هگمپŒن¸¸مپڈمپھم‚‹م€‚مپ¾مپںم€پو™‚é–“مپ®çµŒéپژمپ¨مپ¨م‚‚مپ«ه°ڈمپ•مپھç²’هگمپŒو´—مپ„وµپمپ•م‚Œم€پهگŒمپکم‚µم‚¤م‚؛مپ®ç²’هگمپŒهگŒمپکè·é›¢مپ«م‚ڈمپںمپ£مپ¦ç§»ه‹•مپ—مپ¦مپڈم‚‹مپںم‚پم€پé«که؛¦مپھéپ¸هˆ¥مپŒمپ‹مپ‹م‚‹هڈ¯èƒ½و€§م‚‚مپ‚م‚‹م€‚مپ¾مپںم€په°‘مپھمپڈمپ¨م‚‚ه²©çں³ç²’هگمپ«ç”±و¥مپ™م‚‹ه †ç©چ物مپ§مپ¯م€په †ç©چ物مپŒهڈ¤مپڈوˆگç†ںمپ—مپ¦مپ„م‚‹مپ»مپ©çں³è‹±ï¼ˆم‚¯م‚©مƒ¼مƒ„)مپ®هگ«وœ‰é‡ڈمپŒé«کمپڈمپھم‚‹م€‚م‚¯م‚©مƒ¼مƒ„مپ¯هœ°çگƒمپ®ه²©çں³مپ«م‚ˆمپڈ見م‚‰م‚Œم‚‹é‰±ç‰©مپ§مپ‚م‚ٹم€پéه¸¸مپ«ç،¬مپڈم€پو‘©è€—مپ«ه¼·مپ„م€‚مپمپ®مپںم‚پو™‚é–“مپ®çµŒéپژمپ¨مپ¨م‚‚مپ«م€پن»–مپ®وگو–™مپ§مپ§مپچمپںç²’هگمپŒمپ™م‚ٹو¸›م‚ٹم€پم‚¯م‚©مƒ¼مƒ„مپ مپ‘مپŒو®‹م‚‹م€‚وµœمپ®ç ‚مپ¯éه¸¸مپ«وˆگç†ںمپ—مپںه †ç©چ物مپ§مپ‚م‚ٹم€پن¸»مپ«çں³è‹±مپ§و§‹وˆگمپ•م‚Œمپ¦مپٹم‚ٹم€پç²’هگمپ¯ن¸¸مپ؟م‚’ه¸¯مپ³مپ¦مپٹم‚ٹم€پهگŒو§کمپ®م‚µم‚¤م‚؛ (é«که؛¦مپ«éپ¸هˆ¥مپ•م‚Œمپ¦مپ„م‚‹)مپ§مپ‚م‚‹[1]م€‚ èµ·و؛گوµ·ه؛•ه †ç©چ物مپ¯م€پمپمپ®èµ·و؛گمپ«م‚ˆمپ£مپ¦م‚‚هˆ†é،مپ™م‚‹مپ“مپ¨مپŒمپ§مپچم‚‹م€‚و¬،مپ® 4 مپ¤مپ®م‚؟م‚¤مƒ—مپŒوŒ™مپ’م‚‰م‚Œم‚‹[3][1]م€‚
陸ç”ںه †ç©چ物陸ç”ںه †ç©چ物مپ¯م€پن¸»مپ«وµ·مپ«وµپم‚Œè¾¼م‚“مپ و—¢هکمپ®ه²©çں³مپ®ه°ڈمپ•مپھç ´ç‰‡مپ§و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹م€‚مپ“م‚Œم‚‰مپ®ه †ç©چ物مپ«مپ¯م€په¾®è¦–çڑ„مپھç²کهœںمپ‹م‚‰ه¤§مپچمپھه²©مپ¾مپ§م€پمپ‚م‚‰م‚†م‚‹ç¯„ه›²مپ®ç²’هگم‚µم‚¤م‚؛مپŒهگ«مپ¾م‚Œمپ¦مپ„م‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚ٹم€په¤§هچٹمپ®وµ·ه؛•مپ§è¦‹م‚‰م‚Œم‚‹م€‚ه²©çں³ه †ç©چ物مپ¯م€پ風雨م€پو°´وµپم€پو¸©ه؛¦م‚„و°·çµگمپ«م‚ˆم‚‹ن؛€è£‚م€پمپٹم‚ˆمپ³مپمپ®ن»–مپ®ن¾µé£ںمƒ—مƒم‚»م‚¹مپ®ن½œç”¨مپ«م‚ˆم‚ٹم€په²©çں³م‚„鉱物مپŒم‚ˆم‚ٹه°ڈمپ•مپھç²’هگمپ«هˆ†è§£مپ•م‚Œم‚‹é¢¨هŒ–مپ®مƒ—مƒم‚»م‚¹م‚’é€ڑمپکمپ¦م€پ陸ن¸ٹمپ§ن½œوˆگمپ•م‚Œم‚‹م€‚مپ“م‚Œم‚‰مپ®ن¾µé£ںمپ•م‚Œمپںه°ڈمپ•مپھç²’هگمپ¯م€پمپ•مپ¾مپ–مپ¾مپھمƒ،م‚«مƒ‹م‚؛مƒ م‚’é€ڑمپکمپ¦وµ·مپ¸مپ¨éپ‹مپ°م‚Œم‚‹[1]م€‚
  
ه²©وˆگه †ç©چ物مپ¯é€ڑه¸¸م€پمپم‚Œم‚‰مپŒç”±و¥مپ™م‚‹ه…ƒم€…مپ®ç‰©è³ھمپ®çµ„وˆگم‚’هڈچوک مپ—مپ¦مپ„م‚‹م€پمپمپ®مپںم‚پم€پçں³è‹±م€پé•·çں³م€پç²کهœں鉱物م€پé…¸هŒ–鉄م€پمپٹم‚ˆمپ³é™¸ç”ںوœ‰و©ں物مپھمپ©مپ®م€پهœ°çگƒمپ®ه²©çں³م‚’و§‹وˆگمپ™م‚‹ن¸»è¦پمپھ鉱物مپ«م‚ˆمپ£مپ¦و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹م€‚çں³è‹±(م‚¬مƒ©م‚¹مپ®ن¸»وˆگهˆ†مپ§مپ‚م‚‹ن؛Œé…¸هŒ–م‚±م‚¤ç´ ) مپ¯م€پمپ»مپ¼مپ™مپ¹مپ¦مپ®ه²©çں³مپ«è¦‹م‚‰م‚Œم‚‹وœ€م‚‚ن¸€èˆ¬çڑ„مپھ鉱物مپ® 1 مپ¤مپ§مپ‚م‚ٹم€پو‘©è€—مپ«éه¸¸مپ«ه¼·مپ„مپںم‚پم€پç ‚م‚’هگ«م‚€ه²©çں³ه †ç©چ物مپ®ن¸»è¦پمپھوˆگهˆ†مپ§مپ‚م‚‹[1]م€‚ ç”ں物起و؛گه †ç©چ物ç”ں物起و؛گه †ç©چ物مپ¯م€پç”ں物مپŒو»م‚“مپ مپ¨مپچمپ«و²ˆé™چمپ™م‚‹ç”ں物مپ®و®‹éھ¸مپ«ç”±و¥مپ™م‚‹م€‚ه †ç©چ物مپ«ه¯„ن¸ژمپ™م‚‹مپ®مپ¯ç”ں物مپ®م€Œç،¬مپ„部هˆ†م€چمپ§مپ‚م‚ٹم€پن¾‹مپˆمپ°è²و®»م€پو¯م€پéھ¨و ¼è¦پç´ مپھمپ©مپ§مپ‚م‚‹م€‚مپ“م‚Œم‚‰مپ®éƒ¨هˆ†مپ¯é€ڑه¸¸مƒںمƒچمƒ©مƒ«هŒ–مپ•م‚Œمپ¦مپٹم‚ٹم€پو»ه¾Œمپ«و€¥é€ںمپ«هٹ£هŒ–مپ™م‚‹è‚‰è³ھمپ®م€Œوں”م‚‰مپ‹مپ„部هˆ†م€چم‚ˆم‚ٹم‚‚è…گو•—مپ—مپ«مپڈمپ„[1]م€‚ ه·¨è¦–çڑ„مپھç”ں物起و؛گه †ç©چ物م‚‚مپ®مپ¨مپ—مپ¦مپ¯م€په¤§ه‹ç”ں物مپ®éھ¨و ¼م‚„و¯م€پو®»مپھمپ©مپ®éپ؛物م‚‚هگ«م‚€مپ¨è€ƒمپˆم‚‹مپ“مپ¨مپŒمپ§مپچم‚‹م€‚مپ“مپ®م‚؟م‚¤مƒ—مپ®ه †ç©چ物مپ¯م€پوµ·و´‹مپ®ه¤§éƒ¨هˆ†مپ§مپ‹مپھم‚ٹمپ¾م‚Œمپ§مپ‚م‚‹م€‚مپھمپœمپھم‚‰م€پمپ“مپ®م‚ˆمپ†مپھه¤§ه‹ç”ں物مپ¯ن¸€èˆ¬مپ«م€پمپ“م‚Œم‚‰مپ®و®‹éھ¸مپŒه †ç©چمپ—è“„ç©چمپ™م‚‹مپ®مپ«هچپهˆ†مپھه¯†ه؛¦مپ§و»مپ¬مپ“مپ¨مپ¯مپھمپ„مپ‹م‚‰مپ§مپ‚م‚‹م€‚مپںمپ مپ—م€پ1 مپ¤مپ®ن¾‹ه¤–مپ¨مپ—مپ¦م€پم‚µمƒ³م‚´ç¤پمپŒوŒ™مپ’م‚‰م‚Œم‚‹م€‚مپ“مپ®ç’°ه¢ƒمپ§م€پمپمپ®éپ؛ن½“(مپ™مپھم‚ڈمپ،م‚µمƒ³م‚´مپ®éھ¨و ¼ï¼‰م‚’و®‹مپ™ç”ں物مپŒéه¸¸مپ«è±ٹه¯Œمپ«هکهœ¨مپ—مپ¦مپٹم‚ٹم€په®ںéڑ›مپ«ç†±ه¸¯مپ®ç ‚مپ®ه¤§éƒ¨هˆ†م‚’هچ م‚پم‚‹وˆگهˆ†مپ§م‚‚مپ‚م‚‹[1]م€‚ ه¾®è¦–çڑ„مپھç”ں物起و؛گه †ç©چ物مپ¯م€په¾®ç”ں物مپ®ç،¬مپ„部هˆ†م€پ特مپ«مپمپ®و®»ï¼ˆمƒ†م‚¹مƒˆï¼‰مپ§و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹م€‚ن¸€مپ¤ن¸€مپ¤مپ¯éه¸¸مپ«ه°ڈمپ•مپ„م‚‚مپ®مپ®م€پمپ“م‚Œم‚‰مپ®ç”ں物مپ¯éه¸¸مپ«è±ٹه¯Œمپ«هکهœ¨مپ—م€پو¯ژو—¥و•°هچپه„„ه€‹ن½“م‚‚و»م‚“مپ§مپ„مپڈمپںم‚پم€په¤§é‡ڈمپ®و®»مپŒوµ·ه؛•مپ¸مپ¨و²ˆمپ؟ç”ں物起و؛گمپ®ه †ç©چ物م‚’ن½œم‚ٹه‡؛مپ—مپ¦مپ„م‚‹م€‚ه¾®è¦–çڑ„مپھو®»مپ§و§‹وˆگمپ•م‚Œمپںه †ç©چ物مپ¯م€په·¨è¦–çڑ„مپھç²’هگمپ«م‚ˆم‚‹ه †ç©چ物م‚ˆم‚ٹم‚‚مپ¯م‚‹مپ‹مپ«è±ٹه¯Œمپ§مپ‚م‚ٹم€پمپمپ®م‚µم‚¤م‚؛مپ®ه°ڈمپ•مپ•مپ®مپںم‚پم€پمپچم‚پمپ®ç´°مپ‹مپ„مپ©م‚چمپ©م‚چمپ—مپںه †ç©چ物ه±¤م‚’ن½œم‚ٹه‡؛مپ™م€‚ه †ç©چه±¤مپŒه°‘مپھمپڈمپ¨م‚‚30%مپ®ه¾®è¦–çڑ„مپھç”ں物起و؛گ物è³ھمپ§و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹ه ´هگˆم€پمپم‚Œمپ¯ç”ں物起و؛گمپ®è»ںو³¥مپ¨مپ—مپ¦هˆ†é،مپ•م‚Œم‚‹م€‚مپ“مپ®م‚ˆمپ†مپھه †ç©چ物مپ®و®‹م‚ٹمپ®éƒ¨هˆ†مپ¯م€په¤ڑمپڈمپ®ه ´هگˆم€پç²کهœںمپ§و§‹وˆگمپ•م‚Œم‚‹[1]م€‚ ه †ç©چ物مپ®è§£وگمپ«م‚ˆم‚‹هڈ¤و°—ه€™مپ®وژ¨ه®ڑ
ç”ں物起و؛گمپ®ه †ç©چ物مپ¯م€پé…¸ç´ هگŒن½چن½“و¯”مپ‹م‚‰éپژهژ»مپ®و°—ه€™هڈ²م‚’وژ¨و¸¬مپ™م‚‹مپ“مپ¨مپŒمپ§مپچم‚‹مپںم‚پé‡چè¦پمپ§مپ‚م‚‹م€‚é…¸ç´ هژںهگمپ¯م€پوµ·و°´ن¸مپ« 3 مپ¤مپ®ه½¢و…‹ï¼ˆهگŒن½چن½“)م€پمپ™مپھم‚ڈمپ، O16م€پO17م€پ O18 )مپ§هکهœ¨مپ™م‚‹م€‚O16 مپŒوœ€م‚‚ن¸€èˆ¬çڑ„مپھه½¢و…‹مپ§مپ‚م‚ٹم€پO18مپŒمپ“م‚Œمپ«ç¶ڑمپچم€پO17مپ¯مپ¾م‚Œمپ§مپ‚م‚‹م€‚O16مپ¯O18م‚ˆم‚ٹم‚‚軽مپ„مپںم‚پè’¸ç™؛مپ—م‚„مپ™مپڈم€پO16مپ®ه‰²هگˆمپŒé«کمپ„و°´è’¸و°—مپŒç™؛ç”ںمپ™م‚‹م€‚م‚ˆم‚ٹو¶¼مپ—مپ„و°—ه€™مپ®و™‚وœںمپ«مپ¯م€پو°´è’¸و°—مپŒه‡ç¸®مپ—مپ¦é›¨م‚„é›ھمپ«مپھم‚ٹم€پO16مپ®ه‰²هگˆمپŒé«کمپ„و°·و²³و°·مپŒه½¢وˆگمپ•م‚Œم‚‹م€‚逆مپ«م€پوµ·و°´مپ¯O18 مپ®ه‰²هگˆمپŒو¯”較çڑ„é«کمپڈمپھم‚‹م€‚مپ¾مپںم€پو؛¶هکé…¸ç´ م‚’ç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ¨مپ—مپ¦و®»مپ«çµ„مپ؟è¾¼م‚€وµ·و´‹ç”ں物مپ¯م€پO18هگŒن½چن½“مپ®ه‰²هگˆمپŒé«کمپ„و®»م‚’وŒپمپ¤م€‚مپ“م‚Œمپ¯م€پو®»مپ® O16:O18مپ®و¯”çژ‡مپŒم€په¯’ه†·مپھو°—ه€™مپ®و™‚ن»£مپ«مپ¯ن½ژمپڈم€پو¸©وڑ–مپھو™‚ن»£مپ«مپ¯é«کمپڈمپھم‚‹مپ“مپ¨م‚’و„ڈه‘³مپ™م‚‹م€‚مپھمپœمپھم‚‰م€پو°—ه€™مپŒو¸©وڑ–هŒ–مپ™م‚‹مپ¨و°·و²³مپŒو؛¶مپ‘مپ¦و°·مپ‹م‚‰O16مپŒو”¾ه‡؛مپ•م‚Œمپ¦وµ·مپ«وˆ»م‚ٹم€پو°´ن¸مپ®O16:O18و¯”مپŒه¢—هٹ مپ—م€پç”ں物مپ¯é…¸ç´ م‚’و®»مپ«çµ„مپ؟è¾¼م‚€مپ®مپ§م€پو®»مپ¯م‚ˆم‚ٹé«کمپ„O16:O18و¯”م‚’ç¤؛مپ™مپ“مپ¨مپ«مپھم‚‹مپںم‚پمپ§مپ‚م‚‹م€‚مپ—مپںمپŒمپ£مپ¦م€پ科ه¦è€…مپ¯ç”ں物起و؛گمپ®ه †ç©چ物م‚’èھ؟مپ¹م€پهگŒو™‚مپ«ه¹´ن»£مپŒو—¢çں¥مپ®م‚µمƒ³مƒ—مƒ«م‚’هˆ©ç”¨مپ—مپ¦O16:O18و¯”çژ‡م‚’計算مپ—م€پمپم‚Œم‚‰م‚’و¯”較مپ™م‚‹مپ“مپ¨مپ§م€پمپم‚Œم‚‰مپ®è²و®»مپŒه½¢وˆگمپ•م‚Œمپںو™‚ن»£مپ®و°—ه€™و،ن»¶م‚’وژ¨و¸¬مپ™م‚‹مپ“مپ¨مپŒمپ§مپچم‚‹م€‚هگŒو§کمپ®و¸¬ه®ڑمپ¯م€پو°·مپ®م‚³م‚¢م‚’هˆ©ç”¨مپ—مپ¦م‚‚è،Œمپ†مپ“مپ¨مپŒمپ§مپچم‚‹م€‚ن¸€èˆ¬مپ«م€پو°·مپ®م‚µمƒ³مƒ—مƒ«مپ§O18مپŒ1ppm و¸›ه°‘مپ™م‚‹مپ¨م€پو¸©ه؛¦مپŒ 1.5آ°Cن½ژن¸‹مپ—مپ¦مپ„مپںمپ¨وژ¨و¸¬مپ™م‚‹مپ“مپ¨مپŒمپ§مپچم‚‹[1]م€‚
ه¾®è¦–çڑ„مپھç”ں物起و؛گمپ®ه †ç©چ物مپ®ن¸»مپھن¾›çµ¦و؛گمپ¯م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ (CaCO3) مپ¾مپںمپ¯م‚·مƒھم‚«(SiO2) مپ®و®»م‚’ن½œم‚ٹه‡؛مپ™هچکç´°èƒè—»é،مپٹم‚ˆمپ³هژںç”ںه‹•ç‰©مپ§مپ‚م‚‹م€‚م‚·مƒھم‚«و®»مپ¯م€پçڈھè—»(è—»é،) مپ¨و”¾و•£è™«(هژںç”ںه‹•ç‰©) مپ® 2 مپ¤مپ®ن¸»è¦پمپھم‚°مƒ«مƒ¼مƒ—مپ«ç”±و¥مپ™م‚‹[1]م€‚ çڈھè—»مپ¯م€په…‰هگˆوˆگم‚’è،Œمپ†و¤چ物مƒ—مƒ©مƒ³م‚¯مƒˆمƒ³ï¼ˆه°ڈمپ•مپھو¼‚وµپè—»é،)مپ®é‡چè¦پمپھمƒ،مƒ³مƒگمƒ¼مپ§مپ‚م‚‹م€‚çڈھè—»مپ¯م€پمپم‚Œè‡ھن½“مپŒهˆ†و³Œمپ™م‚‹ç²¾ه·§مپھم‚·مƒھم‚«و®»مپ«ه›²مپ¾م‚Œمپںهچکن¸€مپ®è—»é،ç´°èƒمپ‹م‚‰و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹م€‚çڈھè—»مپ«مپ¯ه¤ڑو§کمپھه½¢çٹ¶مپ®م‚‚مپ®مپŒهکهœ¨مپ—م€پç´°é•·مپ„ç¾½çٹ¶مپ®ه½¢مپ‹م‚‰ه††ه½¢م€پوکںه‹مپھمپ©مپŒمپ‚م‚‹م€‚çڈھè—»مپŒè±ٹه¯Œمپھهœ°هںںمپ§مپ¯م€پن¸‹ه±¤مپ®ه †ç©چ物مپ¯م‚·مƒھم‚«çڈھè—»و®»مپ«ه¯Œم‚“مپ§مپٹم‚ٹم€پمپ“م‚Œمپ¯çڈھè—»هœںمپ¨ه‘¼مپ°م‚Œمپ¦مپ„م‚‹[1]م€‚ و”¾و•£è™«مپ¯ه‹•ç‰©مƒ—مƒ©مƒ³م‚¯مƒˆمƒ³ (مƒ—مƒ©مƒ³م‚¯مƒˆمƒ³و€§هژںç”ںه‹•ç‰©) مپ§مپ‚م‚ٹم€پçڈھè—»مپ¨هگŒو§کمپ«م‚·مƒھم‚«و®»م‚’هˆ†و³Œمپ™م‚‹م€‚و®»مپ¯ç´°èƒم‚’ه›²م‚“مپ§مپ„م‚‹مپŒم€پم‚¢مƒ،مƒ¼مƒگمپ®م‚ˆمپ†مپھ腕(هپ½è¶³ï¼‰م‚’ن¼¸مپ°مپ™مپ“مپ¨مپŒمپ§مپچم‚‹م‚ˆمپ†مپ«ه°ڈمپ•مپھé–‹هڈ£éƒ¨مپ®هگ«م‚“مپ§مپ„م‚‹م€‚و”¾و•£è™«مپ®و®»مپ§مپ¯ه¤ڑمپڈمپ®ه ´هگˆم€پوµ®هٹ›م‚’هٹ©مپ‘م‚‹م‚ˆمپ†مپ«م€پو®»مپ‹م‚‰çھپمپچه‡؛مپںه¤ڑو•°مپ®و£کمپŒè¦‹م‚‰م‚Œم‚‹م€‚çڈھè—»مپ¾مپںمپ¯و”¾و•£è™«مپ®و®»مپŒه¤ڑمپڈم‚’هچ م‚پم‚‹è»ںو³¥مپ¯م€پçڈھè³ھè»ںو³¥مپ¨ه‘¼مپ°م‚Œم‚‹[1]م€‚ çڈھè³ھه †ç©چ物مپ¨هگŒو§کمپ«م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ م€پمپ¤مپ¾م‚ٹçں³çپ°è³ھمپ®ه †ç©چ物م‚‚م€په¾®è¦–çڑ„مپھè—»é،مپٹم‚ˆمپ³هژںç”ںه‹•ç‰©مپ®و®»مپ‹م‚‰ن½œم‚‰م‚Œم‚‹م€‚مپ“مپ®ه ´هگˆم€پن¸»مپھç”±و¥ç”ں物مپ¯ه††çں³è—»ï¼ˆCoccolithophores)مپ¨وœ‰ه”虫مپ§مپ‚م‚‹م€‚ه††çں³è—»مپ¯م€پçڈھè—»م‚ˆم‚ٹم‚‚ç´„100ه€چه°ڈمپ•مپ„هچکç´°èƒمپ®وµ®éپٹو€§è—»é،مپ§مپ‚م‚‹م€‚ه½¼م‚‰مپ®و®»مپ¯م€پç´°èƒم‚’هڈ–م‚ٹه›²م‚€çگƒم‚’ه½¢وˆگمپ™م‚‹ه¤ڑو•°مپ®é€£çµگمپ—مپں CaCO3مƒ—مƒ¬مƒ¼مƒˆ (coccoliths) مپ‹م‚‰و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹م€‚ه††çں³è—»مپŒو»مپ¬مپ¨م€په€‹م€…مپ®مƒ—مƒ¬مƒ¼مƒˆمپŒو²ˆمپ؟م€پè»ںو³¥م‚’ه½¢وˆگمپ™م‚‹م€‚و™‚é–“مپ®çµŒéپژمپ¨مپ¨م‚‚مپ«م€په††çں³è—»مپ®è»ںو³¥مپ¯çں³هŒ–مپ—مپ¦مƒپمƒ§مƒ¼م‚¯مپ«مپھم‚‹م€‚م‚¤مƒ³م‚°مƒ©مƒ³مƒ‰مپ®مƒ›مƒ¯م‚¤مƒˆم‚¯مƒھمƒ•مƒ‰مƒ¼مƒگمƒ¼ï¼ˆWhite Cliffs of Doverم€پمƒ‰مƒ¼مƒگمƒ¼مپ®ç™½مپ„ه´–)مپ¯م€په††çں³è—»مپŒè±ٹه¯Œمپھè»ںو³¥مپ§و§‹وˆگمپ•م‚Œمپں白ن؛œوœںمپ®ه †ç©چ物مپ§مپ‚م‚‹[1]م€‚ وœ‰ه”虫مپ¯م€پم‚«م‚؟مƒ„مƒ مƒھمپ®و®»مپ¨هگŒو§کمپ«م€پ部ه±‹ï¼ˆمƒپمƒ£مƒ³مƒگمƒ¼ï¼‰و§‹é€ مپ®و®»م‚’وŒپمپ¤هژںç”ںه‹•ç‰©مپ§مپ‚م‚‹م€‚ç”ں物مپŒوˆگé•·مپ™م‚‹مپ«مپ¤م‚Œمپ¦م€پè‡ھè؛«مپŒه…¥م‚ٹè¾¼م‚€و–°مپ—مپ„ه¤§مپچمپھ部ه±‹مپŒو§‹ç¯‰مپ•م‚Œم‚‹م€‚مپ»مپ¨م‚“مپ©مپ®وœ‰ه”虫مپ¯ه؛•ç”ںç”ں物مپ§مپ‚م‚ٹم€په †ç©چ物مپ®ن¸ٹم‚„ن¸مپ«ç”ںوپ¯مپ—مپ¦مپ„م‚‹مپŒم€پن¸€éƒ¨مپ«مپ¯و°´وں±مپ®م‚ˆم‚ٹé«کمپ„ن½چç½®مپ«ç”ںوپ¯مپ™م‚‹مƒ—مƒ©مƒ³م‚¯مƒˆمƒ³ç¨®م‚‚هکهœ¨مپ™م‚‹م€‚ه††çں³è—»مپ¨وœ‰ه”虫مپŒو»مپ¬مپ¨م€پمپم‚Œم‚‰مپ¯çں³çپ°è³ھمپ®è»ںو³¥م‚’ه½¢وˆگمپ™م‚‹[1]م€‚ هڈ¤مپ„çں³çپ°è³ھمپ®ه †ç©چه±¤مپ«مپ¯م€پهˆ¥مپ®ç¨®é،مپ®ç”ں物مپ¨مپ—مپ¦Discoastersمپ®و®‹éھ¸م‚‚هگ«مپ¾م‚Œم‚‹مپ“مپ¨مپŒمپ‚م‚‹م€‚مپ“م‚Œمپ¯م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ®و®»م‚’م‚‚مپ¤م€په††çں³è—»مپ¨è؟‘ç¸پمپ®هچکç´°èƒè—»é،مپ§مپ‚م‚‹م€‚Discoasterمپ®و®»مپ¯وکںه½¢مپ§م€پ5ï½40خ¼mمپ®ه¹…م‚µم‚¤م‚؛مپ«éپ”مپ™م‚‹م€‚Discoasterمپ¯ç´„200ن¸‡ه¹´ه‰چمپ«çµ¶و»…مپ—مپںمپŒم€پمپمپ®و®»مپ¯çµ¶و»…ن»¥ه‰چمپ®و™‚ن»£مپ®ه †ç©چ物ه±¤مپŒè¦‹م‚‰م‚Œم‚‹م€پ熱ه¸¯مپ®ه †ç©چ物مپ®و·±éƒ¨مپ«و®‹مپ£مپ¦مپ„م‚‹[1]م€‚ م‚µم‚¤م‚؛مپŒه°ڈمپ•مپ„مپںم‚پم€پمپ“م‚Œم‚‰مپ®و®»مپ¯éه¸¸مپ«م‚†مپ£مپڈم‚ٹمپ¨و²ˆم‚€م€‚ن¸€مپ¤مپ®ه¾®ç”ں物مپ®و®»مپŒوµ·ه؛•مپ¸مپ¨و²ˆم‚€مپ®مپ«م€پç´„10م€œ50ه¹´م‚‚مپ®و™‚é–“مپŒمپ‹مپ‹م‚‹ه ´هگˆم‚‚مپ‚م‚‹م€‚مپمپ®م‚†مپ£مپڈم‚ٹمپ—مپںé™چن¸‹مپ®مپںم‚پم€پ1cm/秒程ه؛¦مپ®ه¼±مپ„وµ·وµپمپ§مپ‚مپ£مپ¦و®»م‚’éپ‹وگ¬مپ™م‚‹مپ«مپ¯هچپهˆ†مپ§مپ‚م‚ٹم€پوµ·ه؛•مپ«هˆ°éپ”مپ™م‚‹مپ¾مپ§مپ«هژں点مپ‹م‚‰ 15,000 kmم‚‚移ه‹•مپ—مپ¦مپ—مپ¾مپ†هڈ¯èƒ½و€§مپŒمپ‚م‚‹م€‚مپ—مپ‹مپ—مپھمپŒم‚‰م€پمپمپ®م‚ˆمپ†مپھé«کمپ„وµپه‹•و€§مپ«م‚‚مپ‹مپ‹م‚ڈم‚‰مپڑم€په †ç©چ物ن¸مپ«هگ«مپ¾م‚Œم‚‹ç”ں物مپ®ç¨®é،مپ¨ç›´ن¸ٹمپ®وµ·و´‹è،¨ه±¤مپ§è¦‹م‚‰م‚Œم‚‹ç”ں物مپ®ç”ں産و€§مپ®ç¨‹ه؛¦مپ¯م€پم‚ˆمپڈن¸€è‡´مپ—مپ¦مپ„م‚‹مپ“مپ¨مپŒه¤ڑمپ„م€‚مپ“مپ®مپ“مپ¨مپ¯م€په †ç©چ物粒هگمپŒمپ¯م‚‹مپ‹مپ«é€ںمپ„é€ںه؛¦مپ§ه؛•مپ«و²ˆمپ؟م€پوµپم‚ŒمپŒمپم‚Œم‚‰م‚’هˆ†و•£مپ•مپ›م‚‹ه‰چمپ«ه †ç©چ物مپ¨مپ—مپ¦ه…ƒمپ®ه ´و‰€مپ®ç›´ن¸‹مپ«è“„ç©چمپ—مپ¦مپ„مپڈمپ“مپ¨م‚’و„ڈه‘³مپ—مپ¦مپ„م‚‹م€‚مپ“مپ®çگ†ç”±مپ¯م€پو®»مپ®مپ»مپ¨م‚“مپ©مپ¯ه®ںéڑ›مپ«مپ¯ه€‹م€…مپ®ç²’هگمپ¨مپ—مپ¦مپ¯و²ˆمپ¾مپڑم€پç´„ 99% مپ¯ن»–مپ®ç”ں物مپ«م‚ˆمپ£مپ¦و¶ˆè²»مپ•م‚Œم€په‡é›†مپ—مپ¦ه¤§مپچمپھç³ç²’مپ¨مپ—مپ¦وژ’ه‡؛مپ•م‚Œم‚‹مپںم‚پمپ§مپ‚م‚‹م€‚مپ“م‚Œمپ¯و®»هچکن½“م‚ˆم‚ٹم‚‚مپ¯م‚‹مپ‹مپ«é€ںمپڈو²ˆمپ؟م€پم‚ڈمپڑمپ‹10ï½15و—¥ç¨‹ه؛¦مپ§وµ·ه؛•مپ¸مپ¨هˆ°éپ”مپ™م‚‹م€‚مپ“م‚Œمپ¯ç²’هگمپŒهˆ†و•£مپ™م‚‹مپ®مپ«هچپهˆ†مپھو™‚é–“م‚’ن¸ژمپˆمپڑم€پن¸‹مپ®ه †ç©چ物مپ¯è،¨é¢è؟‘مپڈمپ§ç™؛ç”ںمپ™م‚‹ç”ں産م‚’هڈچوک مپ™م‚‹م‚‚مپ®مپ§مپ‚م‚‹م€‚مپ“مپ®مƒ،م‚«مƒ‹م‚؛مƒ مپ«م‚ˆم‚‹و²ˆن¸‹é€ںه؛¦مپ®ه¢—هٹ مپ¯م€پç³ن¾؟و€¥è،Œï¼ˆfecal express)مپ¨ه‘¼مپ°م‚Œمپ¦مپ„م‚‹[1]م€‚ وµ·و°´و؛گه †ç©چ物  وµ·و°´مپ«مپ¯مپ•مپ¾مپ–مپ¾مپھو؛¶هک物è³ھمپŒهگ«مپ¾م‚Œمپ¦مپٹم‚ٹم€پهŒ–ه¦هڈچه؟œمپ«م‚ˆمپ£مپ¦مپ“م‚Œم‚‰مپ®ç‰©è³ھمپŒه›؛ن½“ç²’هگمپ¨مپ—مپ¦و²ˆو®؟مپ—م€په †ç©چ物مپ¨مپ—مپ¦è“„ç©چمپ™م‚‹مپ“مپ¨مپŒمپ‚م‚‹م€‚مپ“م‚Œم‚‰مپ®هڈچه؟œمپ¯é€ڑه¸¸م€پو¸©ه؛¦م€پهœ§هٹ›م€پpH مپ®ه¤‰هŒ–مپھمپ©مپ®و،ن»¶مپ®ه¤‰هŒ–مپ«م‚ˆمپ£مپ¦م€پو؛¶è§£çٹ¶و…‹م‚’ç¶وŒپمپ§مپچم‚‹ç‰©è³ھé‡ڈمپŒو¸›ه°‘مپ™م‚‹مپ“مپ¨مپ«م‚ˆم‚ٹه¼•مپچèµ·مپ“مپ•م‚Œم‚‹م€‚ه²©çں³و€§مپ¾مپںمپ¯ç”ں物起و؛گمپ®ه †ç©چ物مپ¨و¯”較مپ—مپ¦م€پوµ·و°´èµ·و؛گمپ®ه †ç©چ物مپ¯ه¤ڑمپڈمپ¯مپھمپ„مپŒم€پ興ه‘³و·±مپ„ه½¢و…‹مپŒمپ„مپڈمپ¤مپ‹هکهœ¨مپ™م‚‹[1]م€‚ ن¾‹مپˆمپ°ç†±و°´ه™´ه‡؛ه”مپ§مپ¯م€پوµ·و°´مپŒوµ·ه؛•مپ«وµ¸é€ڈمپ—م€پمƒم‚°مƒمپ«م‚ˆمپ£مپ¦éپژ熱مپ•م‚Œه™´ه‡؛ه”مپ‹م‚‰و”¾ه‡؛مپ•م‚Œم‚‹م€‚مپ“مپ®éپژ熱و°´مپ«مپ¯ه¤ڑمپڈمپ®و؛¶هک物è³ھمپŒهگ«مپ¾م‚Œمپ¦مپٹم‚ٹم€په™´ه‡؛هڈ£مپ‹م‚‰ه‡؛مپ¦ه†·مپںمپ„وµ·و°´مپ«è§¦م‚Œم‚‹مپ¨م€پن¸»مپ«é‡‘ه±ç،«هŒ–物مپ¨مپ—مپ¦و²ˆو®؟مپ™م‚‹م€‚مپ“م‚Œم‚‰مپ®ç²’هگمپ¯é€ڑو°—هڈ£ï¼ˆمƒپمƒ مƒ‹مƒ¼ï¼‰مپ‹م‚‰وµپم‚Œه‡؛م‚‹م€Œç…™م€چم‚’و§‹وˆگمپ—م€پوœ€çµ‚çڑ„مپ«مپ¯وµ·و°´و؛گه †ç©چ物مپ¨مپ—مپ¦وµ·ه؛•مپ«و²ˆم‚€[1]م€‚熱و°´ه™´ه‡؛ه”مپ¯هœ°çگƒمپ®مƒ—مƒ¬مƒ¼مƒˆه¢ƒç•Œمپ«و²؟مپ£مپ¦هˆ†ه¸ƒمپ—مپ¦مپ„م‚‹مپŒم€پمƒ›مƒƒمƒˆم‚¹مƒمƒƒمƒˆمپ¨ه‘¼مپ°م‚Œم‚‹çپ«ه±±مپھمپ©م€پن¸€éƒ¨مپ¯مƒ—مƒ¬مƒ¼مƒˆمپ®ه†…هپ´مپ«م‚‚見م‚‰م‚Œم‚‹م€‚çڈ¾هœ¨م€پç´„500 مپ®و´»ه‹•çڑ„مپھوµ·ه؛•ç†±و°´ه™´ه‡؛هںںمپŒçں¥م‚‰م‚Œمپ¦مپٹم‚ٹم€پمپمپ®ç´„هچٹهˆ†مپ¯وµ·ه؛•مپ§ç›´وژ¥çڑ„مپ«è¦³ه¯ںمپ•م‚Œم€پو®‹م‚ٹمپ®هچٹهˆ†مپ¯و°´وں±وŒ‡و¨™م‚„وµ·ه؛•ه †ç©چ物مپ‹م‚‰هکهœ¨مپŒن؛ˆوƒ³مپ•م‚Œمپ¦مپ„م‚‹م‚‚مپ®مپ§مپ‚م‚‹[4]م€‚ مƒمƒ³م‚¬مƒ³مƒژه›£ه،ٹ(مƒمƒ³م‚¬مƒ³مƒژم‚¸مƒ¥مƒ¼مƒ«ï¼‰مپ¯م€پوµ·ه؛•مپ«ه½¢وˆگمپ•م‚Œم‚‹مƒمƒ³م‚¬مƒ³م‚„مپمپ®ن»–مپ®é‡‘ه±مپ®ن¸¸مپ„ه،ٹمپ§مپ‚م‚‹م€‚ن¸€èˆ¬مپ«3ï½10cm程ه؛¦مپ®م‚µم‚¤م‚؛مپ§مپ‚م‚‹مپŒم€پن¸مپ«مپ¯ç›´ه¾„30cmمپ«م‚‚éپ”مپ™م‚‹م‚‚مپ®م‚‚مپ‚م‚‹م€‚مپمپ®ه›£ه،ٹمپ¯م€پçœںçڈ مپ«ن¼¼مپںمƒ—مƒم‚»م‚¹مپ§ه½¢وˆگمپ•م‚Œم‚‹م€‚مپ™مپھم‚ڈمپ،م€پن¸ه؟ƒمپ«و ¸مپ¨مپھم‚‹ç‰©è³ھمپŒمپ‚م‚ٹم€پمپمپ®ه‘¨م‚ٹمپ«هگŒه؟ƒه††çٹ¶مپ®ه±¤مپŒم‚†مپ£مپڈم‚ٹمپ¨ه †ç©چمپ—م€پو™‚é–“مپ®çµŒéپژمپ¨مپ¨م‚‚مپ«ه›£ه،ٹمپŒوˆگé•·مپ—مپ¦مپ„مپڈم€‚ه›£ه،ٹمپ®çµ„وˆگمپ¯م€پمپمپ®وµ·هںںم‚„ه½¢وˆگو،ن»¶مپھمپ©مپ«م‚ˆم‚ٹه¤ڑه°‘ç•°مپھم‚‹مپŒم€پé€ڑه¸¸مپ¯é…¸هŒ–مƒمƒ³م‚¬مƒ³مپ¨é…¸هŒ–鉄مپŒه„ھه‹¢مپ§مپ‚م‚‹م€‚مپ¾مپںم€پéٹ…م€پمƒ‹مƒƒم‚±مƒ«م€پم‚³مƒگمƒ«مƒˆمپھمپ©مپ®é‡‘ه±مپŒه°‘é‡ڈهگ«مپ¾م‚Œمپ¦مپ„م‚‹ه ´هگˆم‚‚مپ‚م‚‹م€‚مƒمƒ³م‚¬مƒ³ه›£ه،ٹمپ®و²ˆو®؟مپ¯م€پن»ٹو—¥çں¥م‚‰م‚Œمپ¦مپ„م‚‹وœ€م‚‚éپ…مپ„هœ°è³ھه¦çڑ„مƒ—مƒم‚»م‚¹مپ®1مپ¤مپ§مپ‚م‚ٹم€پ百ن¸‡ه¹´مپ”مپ¨مپ«و•°مƒںمƒھمƒ،مƒ¼مƒˆمƒ«مپ¨مپ„مپ†م‚¹مƒ”مƒ¼مƒ‰مپ§وˆگé•·مپ™م‚‹م€‚مپمپ®مپںم‚پم€پن»–مپ®ه †ç©چ物مپŒه›£ه،ٹم‚’覆مپ„مپم‚Œن»¥ن¸ٹمپ®وˆگé•·م‚’éک»ه®³مپ™م‚‹مپ“مپ¨مپŒç„،مپ„م‚ˆمپ†مپھم€په²©وˆگمƒ»ç”ں物起و؛گه †ç©چ物مپ®è“„ç©چçژ‡مپŒن½ژمپ„وµ·هںںمپ§مپ®مپ؟ه½¢وˆگمپ•م‚Œم‚‹م€‚مپ—مپںمپŒمپ£مپ¦مƒمƒ³م‚¬مƒ³ه›£ه،ٹمپ¯é€ڑه¸¸م€پوµ·ه؛•مپ«ه¤§é‡ڈمپ«è“„ç©چمپ™م‚‹ه²©çں³èµ·و؛گم‚„ç”ں物起و؛گمپ®ه †ç©چ物وµپه…¥مپŒéه¸¸مپ«ه°‘مپھمپ„م€پ陸هœ°مپ‹م‚‰éپ مپڈ離م‚Œمپںن¸ه¤®وµ·و´‹مپ®é کهںںمپ«é™گه®ڑمپ•م‚Œم‚‹(ه›³ 12.4.2 هڈ³)[1]م€‚ ه›£ه،ٹمپ«مپ¯ه¤ڑمپڈمپ®ه•†و¥çڑ„مپ«ن¾،ه€¤مپ®مپ‚م‚‹é‡‘ه±مپŒهگ«مپ¾م‚Œمپ¦مپ„م‚‹مپںم‚پم€پéپژهژ»و•°هچپه¹´مپ«م‚ڈمپںمپ£مپ¦مƒژم‚¸مƒ¥مƒ¼مƒ«مپ®وژ،وژکمپ«ه¤§مپچمپھé–¢ه؟ƒمپŒه¯„مپ›م‚‰م‚Œمپ¦مپچمپ¦مپ„م‚‹م€‚مپ“م‚Œمپ¾مپ§مپ®مپ¨مپ“م‚چم€پمپ»مپ¨م‚“مپ©مپ®هٹھهٹ›مپ¯وژ¢ç´¢و®µéڑژمپ«مپ¨مپ©مپ¾مپ£مپ¦مپ„م‚‹م€‚و·±وµ·وژ،وژکن؛‹و¥مپ®é«کم‚³م‚¹مƒˆم€پوژ،وژکو¨©مپ«é–¢مپ™م‚‹و”؟و²»çڑ„ه•ڈé،Œم€پمپ“م‚Œم‚‰مپ®ه†چç”ںن¸چهڈ¯èƒ½مپھ資و؛گمپ®وژ،وژکم‚’هڈ–م‚ٹه·»مپڈç’°ه¢ƒه•ڈé،Œمپھمپ©م€په¤ڑمپڈمپ®è¦په› مپŒمƒژم‚¸مƒ¥مƒ¼مƒ«مپ®ه¤§è¦ڈو¨،مپھوژ،وژکم‚’ه¦¨مپ’مپ¦مپ„م‚‹[1]م€‚ è’¸ç™؛ه²©ï¼ˆEvaporites)مپ¯م€پوµ·و°´مپŒè’¸ç™؛مپ™م‚‹مپ¨مپچمپ«ه½¢وˆگمپ•م‚Œم‚‹وµ·و°´èµ·و؛گه †ç©چ物مپ§مپ‚م‚ٹم€پو؛¶è§£مپ—مپں物è³ھمپŒه›؛ن½“م€پ特مپ«ه²©ه،©(ه،©م€پNaCl) مپ¨مپ—مپ¦و²ˆو®؟مپ—مپںم‚‚مپ®مپ§مپ‚م‚‹م€‚وµ·و°´مپ®è’¸ç™؛مپ¯é£ں用ه،©مپ®ç”ں産مپ®وœ€م‚‚هڈ¤ه…¸çڑ„مپھه½¢ه¼ڈمپ§مپ‚م‚ٹم€پن»ٹو—¥مپ§م‚‚è،Œم‚ڈم‚Œمپ¦مپ„م‚‹م‚‚مپ®مپ§مپ‚م‚‹م€‚ه²©ه،©è’¸ç™؛ه²©مپ®ه¤§è¦ڈو¨،مپھه †ç©چ物مپ¯م€پهœ°ن¸وµ·مپ®ن¸‹م‚’هگ«م‚€ه¤ڑمپڈمپ®ه ´و‰€مپ«هکهœ¨مپ™م‚‹م€‚ن¾‹مپˆمپ°م€پç´„600ن¸‡ه¹´ه‰چمپ«ه§‹مپ¾مپ£مپںهœ°و®»ه¤‰ه‹•مپ«م‚ˆمپ£مپ¦م€په¤§è¥؟و´‹مپ‹م‚‰هœ°ن¸وµ·مپŒéپ®و–مپ•م‚Œم€پو¸©وڑ–مپھو°—ه€™مپ«م‚ˆمپ£مپ¦ه¤§é‡ڈمپ®و°´مپŒè’¸ç™؛مپ—مپںمپںم‚پم€پهœ°ن¸وµ·مپ¯مپ»مپ¼ه®Œه…¨مپ«ن¹¾ç‡¥مپ—ه¤§é‡ڈمپ®ه،©مپ®ه †ç©چ物مپŒو®‹مپ•م‚Œمپںم€‚مپ“م‚Œمپ¯Messinian Salinity Crisisمپ¨مپ—مپ¦çں¥م‚‰م‚Œمپ¦مپ„م‚‹م€‚مپ•م‚‰مپ«هœ°ن¸وµ·مپ¯ç´„530ن¸‡ه¹´ه‰چمپ«م‚‚ه†چمپ³و°¾و؟«مپ—م€پمپم‚Œم‚‰مپ®و°¾و؟«مپ«م‚ˆمپ£مپ¦ç”ںمپ¾م‚Œمپںه²©ه،©ه †ç©چ物مپ¯ن»–مپ®ه †ç©چ物مپ«م‚ˆم‚ٹهں‹و²،مپ—مپںمپŒم€پن¸€éƒ¨مپ¯وµ·ه؛•مپ«و®‹هکمپ—مپ¦مپ„م‚‹[1]م€‚  وµ·ه؛•مپ®مƒمƒ³م‚¬مƒ³ه›£ه،ٹ  م‚؟م‚¤مپ§è¦‹م‚‰م‚Œم‚‹وµ·و°´مپ®è’¸ç™؛مپ«م‚ˆم‚‹é£ں用ه،©مپ®ç”ں産  مƒگمƒڈمƒمپ®مƒ“مƒ¼مƒپمپ§è¦‹م‚‰م‚Œم‚‹م‚ھمƒ¼مƒ©م‚¤مƒˆ م‚ھمƒ¼مƒ©م‚¤مƒˆï¼ˆOolites)مپ¯م€پوµ®éپٹç²’هگمپ®ه‘¨م‚ٹمپ®وگو–™مپ®هگŒه؟ƒه††çٹ¶مپ®ه±¤مپ‹م‚‰ه½¢وˆگمپ•م‚Œمپںم€په°ڈمپ•مپڈن¸¸مپ؟م‚’ه¸¯مپ³مپںç²’هگمپ§مپ‚م‚‹م€‚مپم‚Œم‚‰مپ¯é€ڑه¸¸م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ§و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹مپŒم€پمƒھمƒ³é…¸ه،©م‚„مپمپ®ن»–مپ®ç‰©è³ھمپ‹م‚‰و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹ه ´هگˆم‚‚مپ‚م‚‹م€‚م‚ھمƒ¼مƒ©م‚¤مƒˆمپ¯م‚ھمƒ¼مƒ©م‚¤مƒˆç ‚مپ¨مپ—مپ¦è“„ç©چمپ•م‚Œم€پمپ“م‚Œمپ¯مƒگمƒڈمƒمپ§وœ€م‚‚è±ٹه¯Œمپ«è¦‹م‚‰م‚Œم‚‹[1]م€‚ مƒ،م‚؟مƒ³مƒڈم‚¤مƒ‰مƒ¬مƒ¼مƒˆمپ¯م€پ産و¥ç”¨é€”مپ®هڈ¯èƒ½و€§مپŒمپ‚م‚‹م€پوµ·و´‹èµ·و؛گمپ®é‰±ه؛ٹمپ§مپ‚م‚‹م€‚مپ™مپ¹مپ¦مپ®é™¸و؛گç”ںوˆگ物مپ«مپ¯م€پن¸»مپ«é™¸ç”ںو¤چ物مپ«ç”±و¥مپ™م‚‹ه°‘é‡ڈمپ®وœ‰و©ں物مپŒهگ«مپ¾م‚Œمپ¦مپ„م‚‹م€‚مپ“مپ®ه°‘é‡ڈمپ®وœ‰و©ں物مپ«هٹ مپˆم€پوµ·و´‹و¤چ物م‚„ه‹•ç‰©مپ‹م‚‰مپ®وœ‰و©ں物مپŒم€پ特مپ«وµ·ه²¸مپ‹م‚‰و•°ç™¾kmن»¥ه†…مپ®é™¸ç”ںه †ç©چ物مپ¨مپ—مپ¦è“„ç©چمپ™م‚‹م€‚ه †ç©چ物مپŒç©چمپ؟é‡چمپھم‚‹مپ¨م€پم‚ˆم‚ٹو·±مپ„部هˆ†مپŒهœ°ç†±مپ«م‚ˆمپ£مپ¦هٹ 熱مپ•م‚Œم€په¾®ç”ں物مپ®هƒچمپچمپ«م‚ˆم‚ٹوœ‰و©ں物مپŒهˆ†è§£مپ•م‚Œمپ¯مپکم‚پم‚‹م€‚مپ“م‚Œمپ¯é…¸ç´ مپ®مپھمپ„çٹ¶و…‹ (ه«Œو°—و€§و،ن»¶) مپ§è،Œم‚ڈم‚Œم‚‹مپںم‚پم€پمپ“مپ®ن»£è¬مپ®ه‰¯ç”£ç‰©مپ¯مƒ،م‚؟مƒ³م‚¬م‚¹ (CH4) مپ§مپ‚م‚‹م€‚ç´°èڈŒمپ«م‚ˆمپ£مپ¦و”¾ه‡؛مپ•م‚Œمپںمƒ،م‚؟مƒ³مپ¯م€پوµ·ه؛•مپ«هگ‘مپ‹مپ£مپ¦ه †ç©چ物م‚’é€ڑمپ£مپ¦م‚†مپ£مپڈم‚ٹمپ¨ن¸ٹهگ‘مپچمپ«ن¸ٹوک‡مپ™م‚‹م€‚و°´و·±500mï½1,000mمپ®وµ·ه؛•مپ¯ن½ژو¸©ï¼ˆ4℃)مپ§مپ‚م‚ٹم€پو°´مپ¨مƒ،م‚؟مƒ³مپŒçµگهگˆمپ—مپ¦م€پمƒ،م‚؟مƒ³مƒڈم‚¤مƒ‰مƒ¬مƒ¼مƒˆمپ¨مپ—مپ¦çں¥م‚‰م‚Œم‚‹ç‰©è³ھم‚’ç”ںوˆگمپ™م‚‹م€‚وµ·ه؛•ن¸‹مپ®و•°مƒ،مƒ¼مƒˆمƒ«مپ‹م‚‰و•°ç™¾مƒ،مƒ¼مƒˆمƒ«ç¨‹ه؛¦مپ®ه †ç©چ物و·±ه؛¦مپ§مپ¯م€پمƒ،م‚؟مƒ³مƒڈم‚¤مƒ‰مƒ¬مƒ¼مƒˆمپŒه®‰ه®ڑمپ™م‚‹ç¨‹ه؛¦مپ«و¸©ه؛¦مپŒن½ژمپ„مپںم‚پم€پمƒڈم‚¤مƒ‰مƒ¬مƒ¼مƒˆمپ¯ه †ç©چ物ه†…مپ«è“„ç©چمپ•م‚Œم‚‹م€‚مƒ،م‚؟مƒ³مƒڈم‚¤مƒ‰مƒ¬مƒ¼مƒˆمپ¯م€پهٹ 熱مپ™م‚‹مپ¨مƒ،م‚؟مƒ³م‚¬م‚¹مپ¨مپ—مپ¦و”¾ه‡؛مپ•م‚Œم‚‹م€پهڈ¯ç‡ƒو€§مپ®ç‰©è³ھمپ§مپ‚م‚‹م€‚وµ·ه؛•ه †ç©چ物ه†…مپ®مƒ،م‚؟مƒ³مپ¯م€پهŒ–çں³ç‡ƒو–™م‚¨مƒچمƒ«م‚®مƒ¼مپ®è†¨ه¤§مپھ貯蔵ه؛«مپ§مپ‚م‚‹م€‚و§کم€…مپھم‚¨مƒچمƒ«م‚®مƒ¼ن¼پو¥م‚„ه›½ه®¶و”؟ه؛œمپ¯م€پمپ“مپ®مƒ،م‚؟مƒ³م‚’ه›هڈژمپ—مپ¦è²©ه£²مپ™م‚‹و–¹و³•م‚’é–‹ç™؛مپ—م‚ˆمپ†مپ¨مپ—مپ¦مپ„م‚‹[1][2]م€‚ ه®‡ه®™èµ·و؛گه †ç©چ物 ه®‡ه®™èµ·و؛گمپ®ه †ç©چ物مپ¯م€پهœ°çگƒه¤–مپ®ن¾›çµ¦و؛گمپ«ç”±و¥مپ—م€په¾®è¦–çڑ„مپھه°ڈçگƒن½“(Spherules)مپ¨م‚ˆم‚ٹه¤§مپچمپھéڑ•çں³مپ®ç ´ç‰‡مپ¨مپ„مپ†2種é،مپ«ه¤§مپچمپڈهˆ†é،مپ§مپچم‚‹م€‚ه°ڈçگƒن½“مپ¯ن¸»مپ«م‚·مƒھم‚«م‚„鉄م€پمƒ‹مƒƒم‚±مƒ«مپ§و§‹وˆگمپ•م‚Œمپ¦مپٹم‚ٹم€پوµپوکںمپŒه¤§و°—هœڈمپ«çھپه…¥مپ—مپںه¾Œمپ«ç‡ƒمپˆه°½مپچم‚‹éڑ›مپ«و”¾ه‡؛مپ•م‚Œم‚‹مپ¨è€ƒمپˆم‚‰م‚Œمپ¦مپ„م‚‹م€‚éڑ•çں³مپ®ç ´ç‰‡مپ¯م€پéڑ•çں³مپŒهœ°çگƒمپ«è،çھپمپ—مپ¦مپ§مپچمپںم‚‚مپ®مپ§مپ‚م‚‹م€‚مپ“م‚Œم‚‰مپ®ه¼·مپ„è،و’ƒمپ®è،çھپمپ«م‚ˆمپ£مپ¦ç²’هگمپŒه¤§و°—ن¸مپ«و”¾ه‡؛مپ•م‚Œم€پوœ€çµ‚çڑ„مپ«مپ¯هœ°è،¨مپ«وˆ»م‚ٹم€په †ç©چ物مپ¨مپ—مپ¦è“„ç©چمپ•م‚Œم‚‹م€‚ه°ڈçگƒن½“مپ¨هگŒو§کمپ«م€پéڑ•çں³مپ®ç ´ç‰‡مپ¯مپ»مپ¨م‚“مپ©مپŒم‚·مƒھم‚«م‚„鉄م€پمƒ‹مƒƒم‚±مƒ«مپ‹م‚‰وˆگم‚‹م€‚مپ“م‚Œم‚‰مپ®è،çھپمپ«م‚ˆم‚‹ç ´ç‰‡مپ® 1 مپ¤مپ®ه½¢و…‹مپ¯م€پم‚¬مƒ©م‚¹çٹ¶مپ®ه°ڈمپ•مپھو¶²و»´مپ§مپ‚م‚‹مƒ†م‚¯م‚؟م‚¤مƒˆمپ§مپ‚م‚‹م€‚مپم‚Œم‚‰مپ¯م€پéڑ•çں³مپ®è،çھپن¸مپ«و”¾ه‡؛مپ•م‚Œمپ¦و؛¶èچمپ—مپںهœ°çگƒن¸ٹç”±و¥مپ®م‚·مƒھم‚«مپ§و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹هڈ¯èƒ½و€§مپŒé«کمپڈم€پهœ°è،¨مپ«وˆ»مپ£مپںمپ¨مپچمپ«ه†·هچ´مپ•م‚Œه›؛هŒ–مپ™م‚‹مپ“مپ¨مپ§ن½œم‚‰م‚Œم‚‹[1]م€‚ ه®‡ه®™èµ·و؛گمپ®ه †ç©چ物مپ¯وµ·و´‹مپ§مپ¯مپ‹مپھم‚ٹمپ¾م‚Œمپ§مپ‚م‚ٹم€پé€ڑه¸¸م€په †ç©چ物مپ¨مپ—مپ¦ه¤§è¦ڈو¨،مپ«è“„ç©چمپ™م‚‹مپ“مپ¨مپ¯مپھمپ„م€‚مپ—مپ‹مپ—م€پمپم‚Œمپ¯هœ°çگƒمپ«é™چم‚ٹو³¨مپگه®‡ه®™ه،µم‚’é€ڑمپ—مپ¦م€پ絶مپˆمپڑè؟½هٹ مپ•م‚Œمپ¦مپ„م‚‹هکهœ¨مپ§م‚‚مپ‚م‚‹م€‚飛و¥مپ™م‚‹ه®‡ه®™مپ‹م‚‰مپ®م‚´مƒں(éڑ•çں³ç‰ï¼‰مپ®ç´„90% مپ¯ه¤§و°—هœڈمپ«çھپه…¥مپ™م‚‹éڑ›مپ«و°—هŒ–مپ™م‚‹مپŒم€پمپم‚Œمپ§م‚‚و¯ژو—¥5ï½300مƒˆمƒ³مپ®ه®‡ه®™ه،µمپŒهœ°è،¨مپ«ç€é™¸مپ™م‚‹مپ¨وژ¨ه®ڑمپ•م‚Œمپ¦مپ„م‚‹[1]م€‚ و§‹وˆگçڈھè³ھè»ںو³¥
çڈھè³ھè»ںو³¥مپ¯م€پو·±وµ·ه؛•مپ«ن½چç½®مپ™م‚‹ç”ں物起و؛گمپ®éپ و´‹و€§ه †ç©چ物مپ®ن¸€ç¨®مپ§مپ‚م‚‹م€‚çڈھè³ھè»ںو³¥مپ¯و·±وµ·ه †ç©چ物مپ®ن¸مپ§وœ€م‚‚ن¸€èˆ¬çڑ„مپ§مپ¯مپھمپڈم€پوµ·ه؛•مپ®ç´„ 15% م‚’هچ م‚پمپ¦مپ„م‚‹[5]م€‚è»ںو³¥مپ¯م€پéپ و´‹ه¾®ç”ں物مپ®éھ¨و ¼مپ®و®‹éھ¸م‚’ه°‘مپھمپڈمپ¨م‚‚ 30% هگ«م‚€ه †ç©چ物مپ¨ه®ڑ義مپ•م‚Œم‚‹[6]م€‚çڈھè³ھè»ںو³¥مپ¯م€پن¸»مپ«çڈھè—»م‚„و”¾و•£è™«مپھمپ©مپ®ه¾®è¦–çڑ„مپھوµ·و´‹ç”ں物مپ®م‚·مƒھم‚«مƒ™مƒ¼م‚¹مپ®éھ¨و ¼مپ§و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹م€‚ه¤§é™¸ç¸پè¾؛ن»کè؟‘مپ®çڈھè³ھè»ںو³¥مپ®ن»–مپ®وˆگهˆ†مپ«مپ¯م€پ陸ç”ںç”±و¥مپ®م‚·مƒھم‚«ç²’هگمپ¨وµ·ç¶؟éھ¨ç‰‡مپŒهگ«مپ¾م‚Œم‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹م€‚çڈھè³ھè»ںو³¥مپ¯م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ ç”ں物 (مپ™مپھم‚ڈمپ،م€په††çں³è—») مپ®éھ¨و ¼مپ‹م‚‰ن½œم‚‰م‚Œم‚‹çں³çپ°è³ھè»ںو³¥مپ¨مپ¯ه¯¾ç…§çڑ„مپ«م€پم‚ھمƒ‘مƒ¼مƒ« م‚·مƒھم‚«Si(O2)مپ‹م‚‰ن½œم‚‰م‚Œم‚‹éھ¨و ¼مپ§و§‹وˆگمپ•م‚Œم‚‹م€‚م‚·مƒھم‚« (Si) مپ¯ç”ں物مپ«ن¸چهڈ¯و¬ مپھè¦پç´ مپ§مپ‚م‚ٹم€پم‚·مƒھم‚« م‚µم‚¤م‚¯مƒ«م‚’é€ڑمپکمپ¦وµ·و´‹ç’°ه¢ƒمپ§هٹ¹çژ‡çڑ„مپ«مƒھم‚µم‚¤م‚¯مƒ«مپ•م‚Œم‚‹[7]م€‚陸هœ°مپ‹م‚‰مپ®è·é›¢م€پو°´و·±م€پوµ·مپ®è‚¥و²ƒه؛¦مپ¯مپ™مپ¹مپ¦م€پوµ·و°´ن¸مپ®م‚ھمƒ‘مƒ¼مƒ« م‚·مƒھم‚«هگ«وœ‰é‡ڈمپ¨çڈھè³ھè»ںو³¥مپ®هکهœ¨مپ«ه½±éں؟م‚’ن¸ژمپˆم‚‹è¦په› مپ®ن¸€مپ¤مپ§مپ‚م‚‹م€‚
çں³çپ°è³ھمپ®è»ںو³¥و–¹è§£çں³م‚„م‚¢مƒ©م‚´مƒٹم‚¤مƒˆمپ¨مپ„مپ£مپںç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ®ه‰²هگˆمپŒé«کمپ„ه †ç©چ物م‚„ه †ç©چه²©مپ¯م€پçں³çپ°è³ھ(calcareous)ه †ç©چ物مپ¨ه‘¼ç§°مپ•م‚Œم‚‹م€‚çں³çپ°è³ھمپ®ه †ç©چ物 (çں³çپ°ه²©) مپ¯é€ڑه¸¸م€پ陸هœ°è؟‘مپڈمپ®وµ…瀬مپ«ه †ç©چمپ™م‚‹م€‚مپ“م‚Œمپ¯م€پ陸هœ°ç”±و¥مپ®و „é¤ٹç´ م‚’ه؟…è¦پمپ¨مپ™م‚‹وµ·و´‹ç”ں物مپ«م‚ˆمپ£مپ¦ç‚é…¸ه،©مپŒو²ˆو®؟مپ™م‚‹مپںم‚پمپ§مپ‚م‚‹م€‚ن¸€èˆ¬çڑ„مپ«é™¸هœ°مپ‹م‚‰é›¢م‚Œم‚‹مپ»مپ©م€په †ç©چ物ن¸مپ®çں³çپ°è³ھمپ¯ه°‘مپھمپڈمپھم‚‹م€‚مپںمپ مپ—ن¸€éƒ¨مپ®هœ°هںںمپ§مپ¯م€پهµگم‚„وµ·وµپمپ®ه¤‰هŒ–مپ«م‚ˆم‚ٹم€پçں³çپ°è³ھمپ®ه †ç©چ物مپŒه¤ڑمپڈ見م‚‰م‚Œم‚‹ه ´و‰€م‚‚مپ‚م‚‹م€‚çں³çپ°è³ھè»ںو³¥مپ¯م€پوµ·ه؛•مپ«è“„ç©چمپ™م‚‹مƒ—مƒ©مƒ³م‚¯مƒˆمƒ³ç”ں物مپ«ç”±و¥مپ™م‚‹ç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ®ن¸€ç¨®مپ§مپ‚م‚‹م€‚مپ“م‚Œمپ¯م€پوµ·و´‹مپŒç‚é…¸ه،©è£œه„ںو·±ه؛¦م‚ˆم‚ٹم‚‚وµ…مپ„ه ´هگˆمپ«مپ®مپ؟ç™؛ç”ںمپ™م‚‹م€‚مپ“مپ®و·±مپ•م‚ˆم‚ٹن¸‹مپ§مپ¯م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپŒوµ·و´‹و°´مپ«و؛¶è§£مپ—ه§‹م‚پم€پçڈھè³ھè»ںو³¥م‚„éپ و´‹و€§èµ¤è‰²ç²کهœںمپھمپ©مپ®éçں³çپ°è³ھه †ç©چ物مپ®مپ؟مپŒه®‰ه®ڑçڑ„مپ«ه †ç©چمپ™م‚‹مپںم‚پمپ§مپ‚م‚‹م€‚
çں³هŒ–ه †ç©چ物詳細مپ¯ه †ç©چه²©م‚’هڈ‚ç…§م€‚ ه †ç©چ物مپ®هˆ†ه¸ƒ ه †ç©چ物مپŒمپ©مپ“مپ«مپ©مپ®م‚ˆمپ†مپ«è“„ç©چمپ™م‚‹مپ‹مپ¯م€پç™؛ç”ںو؛گمپ‹م‚‰مپ®ç‰©è³ھمپ®é‡ڈم‚„م€پç™؛ç”ںو؛گمپ‹م‚‰مپ®è·é›¢م€په †ç©چ物مپŒè“„ç©چمپ™م‚‹مپ®مپ«ه؟…è¦پمپھو™‚é–“م€په †ç©چ物مپ®ن؟هکه؛¦م€په †ç©چم‚·م‚¹مƒ†مƒ مپ«م‚ˆمپ£مپ¦ن¾›çµ¦مپ•م‚Œم‚‹ن»–مپ®ç¨®é،مپ®ه †ç©چ物مپ®é‡ڈم€پمپھمپ©مپ®و§کم€…مپھه½±éں؟م‚’هڈ—مپ‘م‚‹[1]م€‚ ه †ç©چ物مپ®ه †ç©چé€ںه؛¦مپ¯م€پوµ·و´‹مپ®ه¤§éƒ¨هˆ†مپ§و¯”較çڑ„éپ…مپڈم€په¤ڑمپڈمپ®ه ´هگˆم€پوکژç¢؛مپھه †ç©چ物مپŒه½¢وˆگمپ•م‚Œم‚‹مپ¾مپ§مپ«و•°هچƒه¹´مپ‹مپ‹م‚‹م€‚ه²©è³ھه †ç©چ物مپ¯وœ€م‚‚é€ںمپڈè“„ç©چمپ—م€پم‚ˆم‚ٹç²—مپ„ç²’هگمپ®ه ´هگˆمپ¯1000ه¹´مپ‚مپںم‚ٹ1مƒ،مƒ¼مƒˆمƒ«ن»¥ن¸ٹمپ®م‚ھمƒ¼مƒ€مƒ¼مپ§مپ‚م‚‹م€‚مپںمپ مپ—م€پوµپé‡ڈمپ®ه¤ڑمپ„ه¤§و²³ه·مپ®و²³هڈ£ن»کè؟‘مپ§مپ¯م€په †ç©چé€ںه؛¦مپŒو،پéپ•مپ„مپ«é«کمپڈمپھم‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹[1]م€‚ ç”ں物由و¥مپ®è»ںو³¥مپ¯م€پ1000ه¹´مپ‚مپںم‚ٹç´„1cm مپ®ه‰²هگˆمپ§è“„ç©چمپ—م€په°ڈمپ•مپھç²کهœںç²’هگمپ¯1000ه¹´مپ‚مپںم‚ٹç´„1mm مپ®ه‰²هگˆمپ§و·±وµ·مپ«ه †ç©چمپ™م‚‹م€‚ه‰چè؟°مپ®م‚ˆمپ†مپ«م€پمƒمƒ³م‚¬مƒ³ه›£ه،ٹمپ®è“„ç©چé€ںه؛¦مپ¯éه¸¸مپ«éپ…مپڈم€پ1000ه¹´مپ”مپ¨مپ«0.001mm程ه؛¦مپ§مپ‚م‚‹[1]م€‚
  وµ·و´‹ه †ç©چ物مپ¯م€په¤§é™¸ç¸پè¾؛è؟‘مپڈمپ§وœ€م‚‚هژڑمپڈم€پ10kmمپ®هژڑمپ•م‚’超مپˆم‚‹مپ“مپ¨مپŒمپ‚م‚‹م€‚مپ“م‚Œمپ¯م€په¤§é™¸ç¸پè¾؛è؟‘مپڈمپ®هœ°و®»مپ¯éه¸¸مپ«هڈ¤مپ„مپ“مپ¨مپŒه¤ڑمپڈم€پé•·وœںé–“مپ®ه †ç©چ物مپ®è“„ç©چمپŒهڈ¯èƒ½مپ§مپ‚م‚ٹم€پمپ¾مپںه¤§é™¸مپ‹م‚‰ه¤§é‡ڈمپ®é™¸ç”ںه †ç©چ物مپŒوµپه…¥مپ—مپ¦مپ„م‚‹مپںم‚پمپ§مپ‚م‚‹م€‚و–°مپ—مپ„وµ·و´‹هœ°و®»مپŒه½¢وˆگمپ•م‚Œمپ¦مپ„م‚‹ن¸ه¤®وµ·ه¶؛ç³»ن»کè؟‘مپ§مپ¯م€پè‹¥مپ„هœ°و®»مپ«ه †ç©چمپ™م‚‹و™‚é–“مپŒه°‘مپھمپ„مپںم‚پم€په †ç©چ物مپ¯è–„مپڈمپھم‚‹[1]م€‚ وµ·ه¶؛مپ®ن¸ه¤®éƒ¨مپ‹م‚‰مپ®è·é›¢مپŒه؛ƒمپŒم‚‹مپ«مپ¤م‚Œمپ¦ه †ç©چ物مپ¯و¬،第مپ«هژڑمپڈمپھم‚ٹم€پوµ·ه¶؛軸مپ‹م‚‰مپ®è·é›¢مپŒ1000 km離م‚Œم‚‹مپ”مپ¨مپ«ه †ç©چ物مپŒç´„100-200mه¢—هٹ مپ™م‚‹م€‚وµ·ه؛•و‹،و•£çژ‡مپ¯20-40km/百ن¸‡ه¹´مپ§مپ‚م‚ٹم€پمپ“م‚Œمپ¯ 2500ن¸‡ه¹´مپ‹م‚‰5000ن¸‡ه¹´مپ”مپ¨مپ«ç´„100-200m程ه؛¦مپ®é€ںه؛¦مپ§ه †ç©چمپ™م‚‹مپ“مپ¨م‚’è،¨مپ—مپ¦مپ„م‚‹[1]م€‚ مپ“مپ®è¨کن؛‹مپ®ه†’é مپ«مپ‚م‚‹ه›³مپ¯م€پوµ·ه؛•مپ®ن¸»è¦پمپھ種é،مپ®ه †ç©چ物مپ®هˆ†ه¸ƒم‚’ç¤؛مپ—مپ¦مپ„م‚‹م€‚ه®‡ه®™èµ·و؛گمپ®ه †ç©چ物مپ¯م€پوµ·مپ®مپ©مپ®éƒ¨هˆ†مپ«م‚‚è،Œمپچç€مپڈهڈ¯èƒ½و€§مپŒمپ‚م‚‹مپŒم€پن»–مپ®ç¨®é،مپ®ه †ç©چ物مپ«هœ§ه€’مپ•م‚Œم‚‹مپ»مپ©ه°‘é‡ڈمپ—مپ‹è“„ç©چمپ•م‚Œمپھمپ„مپںم‚پم€پمپ©مپ®ه ´و‰€مپ§م‚‚و”¯é…چçڑ„مپ§مپ¯مپھمپ„م€‚هگŒو§کمپ«م€پوµ·و°´و؛گه †ç©چ物مپ¯ç‰¹ه®ڑمپ®ه ´و‰€مپ§é«کو؟ƒه؛¦مپ«مپھم‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹مپŒم€پهœ°çگƒè¦ڈو¨،مپ§مپ؟م‚‹مپ¨éه¸¸مپ«é™گه®ڑçڑ„مپھهœ°هںںمپ«مپ¨مپ©مپ¾م‚‹م€‚مپمپ®مپںم‚پم€پهœ°çگƒè¦ڈو¨،مپ®ه †ç©چ物مƒ‘م‚؟مƒ¼مƒ³مپ®è°è«–مپ§مپ¯م€په®‡ه®™ç”ںوˆگه †ç©چ物مپ¨وµ·و°´و؛گه †ç©چ物مپ¯مپ»مپ¨م‚“مپ©ç„،視مپ§مپچم‚‹[1]م€‚ 陸هœ°مپ®وµپه‡؛م€پو²³ه·مپ®وµپه‡؛م€پمپٹم‚ˆمپ³مپمپ®ن»–مپ®مƒ—مƒم‚»م‚¹مپ«م‚ˆم‚ٹم€پمپ“م‚Œم‚‰مپ®ç‰©è³ھمپŒه¤§é™¸و£ڑمپ«ه¤§é‡ڈمپ«ه †ç©چمپ™م‚‹مپںم‚پم€په¤§é™¸ç¸پ部è؟‘مپڈمپ§مپ¯ç²—مپ„ه²©وˆگ/陸وˆگه †ç©چ物مپŒو”¯é…چçڑ„مپ§مپ‚م‚‹م€‚مپ“مپ®ه †ç©چ物مپ®ه¤ڑمپڈمپ¯ه¤§é™¸و£ڑمپ®ن¸ٹم‚„è؟‘è¾؛مپ«ç•™مپ¾م‚‹مپŒم€پو؟پوµپمپھمپ©مپ®çڈ¾è±،مپ«م‚ˆم‚ٹه¤§é™¸و–œé¢مپ‹م‚‰و·±وµ·ه؛• (و·±وµ·ه¹³هژں) مپ¾مپ§éپ‹مپ°م‚Œم‚‹مپ“مپ¨م‚‚مپ‚م‚‹م€‚ه²©è³ھه †ç©چ物مپ¯م€پهژڑمپ„و°·مپ®è¦†مپ„مپŒن¸€و¬،ç”ں産م‚’هˆ¶é™گمپ—مپ¦مپ„م‚‹هŒ—و¥µمƒ»هچ—و¥µوµ·هںںمپ«مپٹمپ„مپ¦م‚‚ن¸€èˆ¬çڑ„مپ§مپ‚م‚ٹم€پو°·و²³مپ®ه´©ه£ٹمپŒو°·مپ®ç¸پمپ«و²؟مپ£مپ¦ه †ç©چ物م‚’ه †ç©چمپ•مپ›م‚‹[1]م€‚ مپ“م‚Œم‚‰مپ®ه²©وˆگه †ç©چ物مپ¯م€پç™؛ç”ںو؛گمپ‹م‚‰éپ مپ„وµ·و´‹مپ®ن¸ه¤®éƒ¨مپھمپ©مپ§مپ¯مپ‚مپ¾م‚ٹن¸€èˆ¬çڑ„مپ§مپ¯مپھمپ„م€‚مپںمپ مپ—éه¸¸مپ«ه°ڈمپ•مپھç²کهœںç²’هگمپ¯ن¾‹ه¤–مپ§مپ‚م‚ٹم€پن»¥ن¸‹مپ«èھ¬وکژمپ™م‚‹م‚ˆمپ†مپ«م€پن»–مپ®ه²©è³ھه †ç©چ物مپŒهˆ°éپ”مپ—مپھمپ„é کهںںمپ«è“„ç©چمپ™م‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹[1]م€‚ ç”ں物起و؛گمپ®ه †ç©چ物مپ®هˆ†ه¸ƒمپ¯م€پن»–مپ®ه †ç©چ物مپ«م‚ˆم‚‹ç”ں産م€پو؛¶è§£م€پمپٹم‚ˆمپ³ه¸Œé‡ˆمپ®é€ںه؛¦مپ«ن¾هکمپ™م‚‹م€‚و²؟ه²¸هœ°هںںمپ¯éه¸¸مپ«é«کمپ„ن¸€و¬،ç”ں産م‚’ç¤؛مپ—مپ¦مپ„م‚‹مپںم‚پم€پمپ“م‚Œم‚‰مپ®هœ°هںںمپ§مپ¯è±ٹه¯Œمپھç”ں物起و؛گمپ®ه †ç©چ物مپŒوœںه¾…مپ•م‚Œم‚‹م€‚مپ—مپ‹مپ—م€پç”ں物起و؛گمپ®è»ںو³¥مپ¨è¦‹مپھمپ•م‚Œم‚‹مپ«مپ¯م€پç”ں物起و؛گمپ®وˆگهˆ†مپŒه †ç©چ物ه…¨ن½“مپ®30%م‚’超مپˆمپ¦مپ„م‚‹ه؟…è¦پمپŒمپ‚م‚‹م€‚ç”ں産و€§مپ®é«کمپ„و²؟ه²¸هœ°هںںمپ§مپ•مپˆم€پç”ں物起و؛گمپ®ç‰©è³ھم‚’هœ§ه€’مپ™م‚‹مپ»مپ©ه¤ڑمپڈمپ®ه²©çں³èµ·و؛گمپ®وµپه…¥مپŒمپ‚م‚ٹم€پمپ“مپ®30%مپ®مپ—مپچمپ„ه€¤مپ«éپ”مپ—مپھمپ„مپ“مپ¨م‚‚ه¤ڑمپ„م€‚مپمپ®مپںم‚پم€پو²؟ه²¸هœ°هںںمپ¯ه²©çں³و€§ه †ç©چ物مپŒو”¯é…چçڑ„مپ§مپ‚م‚ٹم€په²©çں³و€§ç‰©è³ھمپ®وµپه…¥مپŒمپ»مپ¨م‚“مپ©مپھمپ„éپ و´‹ç’°ه¢ƒمپ§مپ¯مپکم‚پمپ¦ç”ں物起و؛گه †ç©چ物مپŒم‚ˆمپڈ見م‚‰م‚Œم‚‹م‚ˆمپ†مپ«مپھم‚‹[1]م€‚  ç”ں物起و؛گمپ®ه †ç©چ物مپŒè“„ç©چمپ™م‚‹مپںم‚پمپ«مپ¯م€پمپمپ®ç”ں産é€ںه؛¦مپŒه¾®ç”ں物و®»مپŒو؛¶è§£مپ™م‚‹é€ںه؛¦م‚ˆم‚ٹم‚‚ه¤§مپچمپڈمپھمپ‘م‚Œمپ°مپھم‚‰مپھمپ„م€‚م‚·مƒھم‚«مپ¯وµ·و´‹ه…¨ن½“مپ§ن¸چ飽ه’Œمپ§مپ‚م‚ٹم€پم‚ˆم‚ٹوڑ–مپ‹مپ„و°´مپ¨ن½ژمپ„هœ§هٹ›مپ§م‚ˆم‚ٹه®¹وک“مپ«وµ·و°´مپ«و؛¶مپ‘م‚‹م€‚مپ¤مپ¾م‚ٹم€پو·±وµ·م‚ˆم‚ٹم‚‚هœ°è،¨è؟‘مپڈمپ§م‚ˆم‚ٹé€ںمپڈو؛¶è§£مپ™م‚‹م€‚مپ—مپںمپŒمپ£مپ¦م‚·مƒھم‚«ه †ç©چ物مپ¯م€پو؛¶è§£مپ™م‚‹م‚ˆم‚ٹم‚‚é€ںمپڈè“„ç©چمپ™م‚‹م‚ˆمپ†مپھم€پç”ں産و€§مپŒé«کمپڈه†·و¶¼مپھهœ°هںںمپ«مپ®مپ؟è“„ç©چمپ™م‚‹م€‚مپ“مپ®م‚ˆمپ†مپھوµ·هںںمپ¨مپ—مپ¦مپ¯م€پو „é¤ٹهˆ†مپŒè±ٹه¯Œمپ§و°´و¸©مپŒن½ژمپ„赤éپ“ن»کè؟‘م‚„é«کç·¯ه؛¦مپ®و¹§وک‡هœ°هںںمپŒهگ«مپ¾م‚Œم‚‹[1]م€‚ 赤éپ“ن»کè؟‘مپ§ه½¢وˆگمپ•م‚Œمپںè»ںو³¥مپ¯é€ڑه¸¸م€پو”¾و•£è™«مپŒه„ھه‹¢مپ§مپ‚م‚‹مپŒم€پو¥µهœ°è»ںو³¥مپ§مپ¯çڈھè—»مپŒم‚ˆم‚ٹن¸€èˆ¬çڑ„مپ§مپ‚م‚‹م€‚م‚·مƒھم‚«و®»مپŒوµ·ه؛•مپ«و²ˆمپ؟م€پمپمپ®ه¾Œمپ«هˆ¥مپ®ه †ç©چ物ه±¤مپ§è¦†م‚ڈم‚Œم‚‹مپ¨م€پمپم‚Œم‚‰مپ¯م‚‚مپ¯م‚„و؛¶è§£مپ›مپڑم€په †ç©چ物مپŒè“„ç©چمپ™م‚‹م€‚وµ·ه؛•مپ®ç´„15%مپ¯çڈھè³ھè»ںو³¥مپ§è¦†م‚ڈم‚Œمپ¦مپ„م‚‹[1]م€‚ ç”ں物起و؛گمپ®ç‚é…¸م‚«مƒ«م‚·م‚¦مƒ ه †ç©چ物م‚‚م€په †ç©چ物مپŒè“„ç©چمپ™م‚‹مپںم‚پمپ«و؛¶è§£م‚’超مپˆم‚‹ç”ں産م‚’ه؟…è¦پمپ¨مپ™م‚‹مپŒم€پ関連مپ™م‚‹مƒ—مƒم‚»م‚¹مپ¯م‚·مƒھم‚«مپ®ه ´هگˆمپ¨مپ¯ه°‘مپ—ç•°مپھم‚‹م€‚ç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ¯م€پé…¸و€§مپ®و°´مپ«و؛¶مپ‘م‚„مپ™مپ„م€‚ه†·مپںمپ„وµ·و°´مپ¯و؛¶هکCO2م‚’ه¤ڑمپڈهگ«مپ؟م€پوڑ–مپ‹مپ„و°´م‚ˆم‚ٹم‚‚م‚ڈمپڑمپ‹مپ«é…¸و€§مپ§مپ‚م‚‹م€‚مپمپ®مپںم‚پم€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ®و®»مپ¯م€پوڑ–مپ‹مپ„熱ه¸¯مپ®هœ°è،¨و°´م‚ˆم‚ٹم‚‚م€په†·مپںمپڈمپ¦و·±مپ„و¥µو€§مپ®و°´مپ«و؛¶مپ‘م‚„مپ™مپ„م€‚ن¸،و¥µمپ§مپ¯و°´مپ¯ن¸€و§کمپ«ه†·مپںمپ„مپ®مپ§م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ¯مپ‚م‚‰م‚†م‚‹و·±مپ•مپ§ه®¹وک“مپ«و؛¶è§£مپ—م€پç‚é…¸ه،©مپ®ه †ç©چ物مپ¯è“„ç©چمپ—مپھمپ„م€‚و¸©ه¸¯مپٹم‚ˆمپ³ç†±ه¸¯هœ°هںںمپ§مپ¯م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ¯م‚ˆم‚ٹو·±مپ„و°´مپ«و²ˆم‚€مپ«مپ¤م‚Œمپ¦م€پم‚ˆم‚ٹه®¹وک“مپ«و؛¶è§£مپ™م‚‹م‚ˆمپ†مپ«مپھم‚‹[1]م€‚ ç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپŒè“„ç©چمپ™م‚‹مپ®مپ¨هگŒمپکé€ںمپ•مپ§و؛¶è§£مپ™م‚‹و·±مپ•مپ¯م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ 補ه„ںو·±ه؛¦مپ¾مپںمپ¯و–¹è§£çں³è£œه„ںو·±ه؛¦ï¼ˆcalcite compensation depthï¼›CCD)مپ¨ه‘¼مپ°م‚Œم‚‹م€‚مƒھم‚½م‚¯مƒ©م‚¤مƒ³ï¼ˆlysocline)مپ¯م€پ م‚µمƒ¼مƒ¢م‚¯مƒ©م‚¤مƒ³مپٹم‚ˆمپ³ه،©هˆ†ه±¤مپ¨هگŒمپکم‚ˆمپ†مپ«م€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ®و؛¶è§£é€ںه؛¦مپŒهٹ‡çڑ„مپ«ه¢—هٹ مپ™م‚‹و·±مپ•مپ®مپ“مپ¨مپ§مپ‚م‚‹م€‚CCD م‚ˆم‚ٹوµ…مپ„و°´و·±مپ§مپ¯م€پç‚é…¸ه،©مپ®è“„ç©چمپŒو؛¶è§£é€ںه؛¦م‚’超مپˆم€پç‚é…¸ه،©مپ®ه †ç©چ物مپŒه †ç©چمپ™م‚‹م€‚ CCD م‚ˆم‚ٹم‚‚و·±مپ„é کهںںمپ§مپ¯م€پو؛¶è§£é€ںه؛¦مپŒç”ں産é‡ڈم‚’ن¸ٹه›م‚ٹم€پç‚é…¸ه،©ه †ç©چ物مپ¯è“„ç©چمپ§مپچمپھمپ„م€‚ CCDمپ¯é€ڑه¸¸م€پو°´و·±4-4.5kmمپ®و·±مپ•مپ«مپ‚م‚‹مپŒم€پهœ°è،¨و°´مپŒه†·مپںمپ„و¥µمپ§مپ¯مپ¯م‚‹مپ‹مپ«وµ…مپ„箇و‰€مپ§è¦‹م‚‰م‚Œم‚‹م€‚مپ—مپںمپŒمپ£مپ¦م€پçں³çپ°è³ھمپ®è»ںو³¥مپ¯م€پمپ»مپ¨م‚“مپ©مپ®ه ´هگˆم€پن¸ه¤®وµ·ه¶؛ç³»م‚„وµ·ه±±م‚„هڈ°هœ°مپھمپ©مپ®و·±مپ•ç´„4km وœھو؛€مپ®ç†±ه¸¯مپ¾مپںمپ¯و¸©ه¸¯مپ®وµ·هںںمپ§è¦‹م‚‰م‚Œم‚‹[1]م€‚ CCD مپ¯م€په¤ھه¹³و´‹م‚ˆم‚ٹم‚‚ه¤§è¥؟و´‹مپ®و–¹مپŒو·±مپڈمپھم‚‹م€‚مپ“م‚Œمپ¯م€په¤ھه¹³و´‹مپ«مپ¯ CO2مپŒه¤ڑمپڈهگ«مپ¾م‚Œمپ¦مپ„م‚‹مپںم‚پم€پو°´مپŒم‚ˆم‚ٹé…¸و€§مپ«مپھم‚ٹم€پç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپŒم‚ˆم‚ٹو؛¶مپ‘م‚„مپ™مپڈمپھم‚‹مپ‹م‚‰مپ§مپ‚م‚‹م€‚مپ™مپھم‚ڈمپ،مپ“مپ®مپ“مپ¨مپ¯م€په¤ھه¹³و´‹مپŒم‚ˆم‚ٹو·±مپ„مپ¨مپ„مپ†ن؛‹ه®ںمپ¨مپ¨م‚‚مپ«م€په¤§è¥؟و´‹مپŒه¤ھه¹³و´‹م‚ˆم‚ٹم‚‚ه¤ڑمپڈمپ®çں³çپ°è³ھه †ç©چ物م‚’هگ«م‚€مپ“مپ¨م‚’و„ڈه‘³مپ—مپ¦مپ„م‚‹م€‚ه…¨ن½“مپ¨مپ—مپ¦م€پوµ·ه؛•مپ®ç´„48%مپ¯çں³çپ°è³ھمپ®è»ںو³¥مپ§هچ م‚پم‚‰م‚Œمپ¦مپ„م‚‹[1]م€‚ و·±وµ·ه؛•مپ®و®‹م‚ٹمپ®éƒ¨هˆ† (ç´„38%) مپ¯م€پو·±وµ·ç²کهœںمپŒهچ م‚پمپ¦مپ„م‚‹م€‚مپ“م‚Œمپ¯م€پç²کهœںه½¢وˆگمپŒه¤§è¦ڈو¨،مپ«èµ·مپچمپ¦مپ„م‚‹مپ¨مپ„م‚ˆم‚ٹمپ¯م€پن»–مپ®م‚؟م‚¤مƒ—مپ®ه †ç©چ物مپ®وµپه…¥مپŒه°‘مپھمپ„مپںم‚پمپ«èµ·مپچمپ¦مپ„م‚‹م‚‚مپ®مپ§مپ‚م‚‹م€‚ç²کهœںç²’هگمپ¯ه¤§éƒ¨هˆ†مپŒé™¸ç”ںèµ·و؛گمپ®م‚‚مپ®مپ§مپ‚م‚‹مپŒم€پéه¸¸مپ«ه°ڈمپ•مپ„مپںم‚پم€پ風م‚„وµ·وµپمپ«م‚ˆمپ£مپ¦ه®¹وک“مپ«هˆ†و•£مپ—م€پن»–مپ®ç¨®é،مپ®ه †ç©چ物مپŒهˆ°éپ”مپ§مپچمپھمپ„é کهںںمپ«هˆ°éپ”مپ™م‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹م€‚ه®ںéڑ›مپ«م€پهŒ—ه¤ھه¹³و´‹ن¸ه¤®éƒ¨مپ§مپ¯ç²کهœںمپŒه„ھه‹¢مپ§مپ‚م‚‹م€‚مپ“مپ®هœ°هںںمپ¯م€پç²—مپ„ه²©è³ھه †ç©چ物مپŒهˆ°éپ”مپ™م‚‹مپ«مپ¯é™¸هœ°مپ‹م‚‰éپ مپ™مپژم‚‹م€‚هگŒو™‚مپ«م€پç”ں物起و؛گمپ®ç²’هگم‚’è“„ç©چمپ™م‚‹مپ«مپ¯ç”ں産و€§مپŒهچپهˆ†مپ§مپ¯مپھمپڈم€پمپ¾مپںçں³çپ°è³ھ物è³ھمپŒو؛¶è§£مپ™م‚‹ه‰چمپ«ه؛•مپ«هˆ°éپ”مپ™م‚‹مپ«مپ¯و·±مپ™مپژم‚‹[1]م€‚ ç²کهœںç²’هگمپ¯éه¸¸مپ«م‚†مپ£مپڈم‚ٹمپ¨è“„ç©چمپ™م‚‹مپںم‚پم€پç²کهœںمپŒه„ھه‹¢مپھو·±وµ·ه؛•مپ«مپ¯مƒمƒ³م‚¬مƒ³ه›£ه،ٹمپ®م‚ˆمپ†مپھو°´ç´ م‚’هگ«م‚€ه †ç©چ物مپŒهکهœ¨مپ™م‚‹مپ“مپ¨مپŒمپ—مپ°مپ—مپ°مپ‚م‚‹م€‚م‚‚مپ—ن»®مپ«مپ“مپ“مپ§ن»–مپ®ç¨®é،مپ®ه †ç©چ物مپŒç”ںوˆگمپ•م‚Œمپںه ´هگˆم€پمپم‚Œمپ¯مپ¯م‚‹مپ‹مپ«و€¥é€ںمپ«è“„ç©چمپ—م€په›£ه،ٹمپŒوˆگé•·مپ™م‚‹ه‰چمپ«مپم‚Œم‚‰م‚’هں‹م‚پمپ¦مپ—مپ¾مپ†[1]م€‚ و²؟ه²¸ه †ç©چ物 وµ…瀬مپ®وµ·و´‹ç’°ه¢ƒمپ¯م€پم‚µمƒ³م‚´ç¤پم‚„ه¤§é™¸و£ڑه¤–ç¸پمپھمپ©م€پوµ·ه²¸مپ¨و·±وµ·مپ®é–“مپ®é کهںںمپ«è¦‹م‚‰م‚Œم‚‹م€‚مپ“مپ®ç’°ه¢ƒمپ®و°´مپ¯وµ…مپڈو¾„م‚“مپ§مپٹم‚ٹ[15]م€پمپ•مپ¾مپ–مپ¾مپھه †ç©چو§‹é€ م€پç‚é…¸ه،©ه²©م€پم‚µمƒ³م‚´ç¤پمپ®ه½¢وˆگم‚’هڈ¯èƒ½مپ«مپ—مپ¦مپ„م‚‹م€‚ ه †ç©چ物è‡ھن½“مپ¯çں³çپ°ه²©مپ§و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹مپ“مپ¨مپŒه¤ڑمپڈم€پوµ…مپڈوڑ–مپ‹مپڈç©ڈم‚„مپ‹مپھوµ·هںںمپ§ه®¹وک“مپ«ه½¢وˆگمپ•م‚Œم‚‹م€‚وµ…مپ„وµ·و´‹ç’°ه¢ƒمپ¯م€پçڈھه²©è³ھمپ¾مپںمپ¯ç‚ç´ è³ھمپ®ه †ç©چ物مپ®مپ؟مپ‹م‚‰و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹مپ¨مپ¯é™گم‚‰مپڑم€پمپ¾مپںن¸،者مپŒه…±هکمپ—مپھمپ„ه ´هگˆم‚‚مپ‚م‚‹م€پن¾‹مپˆمپ°م€پç‚ç´ è³ھه †ç©چ物مپ®مپ؟مپ§و§‹وˆگمپ•م‚Œمپںوµ…مپ„وµ·و´‹ç’°ه¢ƒم‚„م€په®Œه…¨مپ«çڈھç •è³ھه †ç©چ物مپ§و§‹وˆگمپ•م‚Œمپںوµ·و´‹ç’°ه¢ƒمپھمپ©م‚‚مپ‚م‚ٹه¾—م‚‹م€‚وµ…مپ„و°´مپ®وµ·ه؛•ه †ç©چ物مپ¯م€پم‚ˆم‚ٹه¤§مپچمپھç²’هگم‚µم‚¤م‚؛مپ§و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹م€‚مپ“م‚Œمپ¯م€په°ڈمپ•مپھç²’هگمپŒم‚ˆم‚ٹو·±مپ„و°´مپ«و´—مپ„وµپمپ•م‚Œم‚‹مپںم‚پمپ§مپ‚م‚‹م€‚ç‚ç´ è³ھه †ç©چ物مپ§و§‹وˆگمپ•م‚Œم‚‹ه †ç©چه²©ه†…مپ«مپ¯م€پè’¸ç™؛鉱物م‚‚هگ«مپ¾م‚Œم‚‹ه ´هگˆمپŒمپ‚م‚‹[16]م€‚ه †ç©چ物ه†…مپ§è¦‹م‚‰م‚Œم‚‹وœ€م‚‚ن¸€èˆ¬çڑ„مپھè’¸ç™؛ه²©é‰±ç‰©مپ¯م€پçں³è†ڈم€پç،¬çں³è†ڈم€پمپٹم‚ˆمپ³ه²©ه،©مپ§مپ‚م‚‹م€‚مپم‚Œم‚‰مپ¯م€پçµگو™¶ه±¤م€په¤ç«‹مپ—مپںçµگو™¶م€پمپ¾مپںمپ¯çµگو™¶مپ®م‚¯مƒ©م‚¹م‚؟مƒ¼مپ¨مپ—مپ¦ç™؛ç”ںمپ™م‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹[16]م€‚ هœ°è³ھه¹´ن»£مپ«é–¢مپ—مپ¦مپ¯م€په †ç©چن¸مپ§è¦‹م‚‰م‚Œم‚‹ï¼ˆم‚¨مƒ“م‚„م‚«مƒ‹مپ®ï¼‰ç”²ç¾…مپ®ç´„ 75% مپŒوµ…مپ„وµ·و´‹ه †ç©چ物مپ§è¦‹م‚‰م‚Œم‚‹مپ“مپ¨مپ‹م‚‰م€پé،•ç”ںن»£مپ®ه †ç©چه²©مپ®مپ»مپ¨م‚“مپ©مپ¯وµ…مپ„وµ·و´‹ç’°ه¢ƒمپ«ه †ç©چمپ—مپںمپ¨è€ƒمپˆم‚‰م‚Œمپ¦مپ„م‚‹م€‚ه…ˆم‚«مƒ³مƒ–مƒھم‚¢وœںمپ®ه †ç©چه²©م‚‚م€پهگŒو§کمپ«وµ…مپ„وµ·هںںمپ«ه †ç©چمپ—مپںمپ¨وƒ³ه®ڑمپ•م‚Œم‚‹[17]م€‚مپ“مپ®ه‚¾هگ‘مپ¯م€پهŒ—ç±³م‚„م‚«مƒھمƒ–هœ°هںںمپ§è¦‹م‚‰م‚Œم‚‹[18]م€‚مپ¾مپںم€پ超ه¤§é™¸مپ®هˆ†è£‚م‚„مپمپ®ن»–مپ®و§‹é€ مƒ—مƒ¬مƒ¼مƒˆمپ®ç§»ه‹•éپژ程مپ®çµگوœمپ¨مپ—مپ¦م€پوµ…مپ„وµ·ه؛•ه †ç©چ物مپ¯م€پهœ°è³ھو™‚ن»£مپ®é‡ڈمپ«é–¢مپ—مپ¦ه¤§مپچمپھه¤‰ه‹•م‚’ç¤؛مپ™[18]م€‚   ç”ں物و“¾ن¹±â†’詳細مپ¯م€Œç”ں物و“¾ن¹±م€چم‚’هڈ‚ç…§
ç”ں物و“¾ن¹±مپ¨مپ¯م€په‹•ç‰©م‚„و¤چ物مپ«م‚ˆم‚‹ه †ç©چ物مپ®ه†چهٹ ه·¥مپ®مƒ—مƒم‚»م‚¹مپ§مپ‚م‚‹م€‚مپ“م‚Œم‚‰مپ«مپ¯م€پهœںç ‚ç²’مپ®ç©´وژکم‚ٹم‚„و‘‚هڈ–م€پوژ’ن¾؟مپŒهگ«مپ¾م‚Œم‚‹م€‚ç”ں物و”ھو‹Œو´»ه‹•مپ¯ç’°ه¢ƒمپ«ه¤§مپچمپھه½±éں؟م‚’ن¸ژمپˆم€پç”ں物ه¤ڑو§کو€§مپ®ن¸»è¦پمپھهژںه‹•هٹ›مپ«مپھم‚‹مپ¨è€ƒمپˆم‚‰م‚Œمپ¦مپ„م‚‹[19][20]م€‚ç”ں物و“¾ن¹±مپ®ç ”究مپ¯م€پ1800ه¹´ن»£مپ«مƒپمƒ£مƒ¼مƒ«م‚؛مƒ»مƒ€مƒ¼م‚¦م‚£مƒ³مپŒه؛مپ§ه®ں験م‚’è،Œمپ†مپ“مپ¨مپ«م‚ˆمپ£مپ¦ه§‹مپ¾مپ£مپں[20]م€‚ç”ں物و”ھو‹Œمپ«م‚ˆم‚‹و°´ç”ںه †ç©چ物مپ¨é™¸هœ°هœںه£Œمپ®ç ´ه£ٹمپ¯م€پé‡چè¦پمپھç”ںو…‹ç³»م‚µمƒ¼مƒ“م‚¹م‚’وڈگن¾›مپ™م‚‹م€‚مپ“م‚Œم‚‰مپ«مپ¯م€پو°´ç”ںه †ç©چ物مپ¨مپمپ®ç›´ن¸ٹو°´مپ®و „é¤ٹç´ مپ®ه¤‰هŒ–م€په·£ç©´مپŒن»–مپ®ç”ں物種مپ«مپ¨مپ£مپ¦مپ®éپ؟難و‰€مپ¨مپ—مپ¦ن½؟م‚ڈم‚Œم‚‹مپ“مپ¨م€پمپٹم‚ˆمپ³é™¸ن¸ٹمپ§مپ®هœںه£Œمپ®ç”ں産مپھمپ©مپŒهگ«مپ¾م‚Œم‚‹[21][22]م€‚ ç”ں物و“¾ن¹±ç¨®ï¼ˆمƒگم‚¤م‚ھم‚؟مƒ¼مƒ™مƒ¼م‚؟مƒ¼ï¼‰مپ¯م€پç’°ه¢ƒمپ«ç‰©çگ†çڑ„مپھه¤‰هŒ–م‚’هٹ مپˆم‚‹مپ“مپ¨مپ§ن»–مپ®ç¨®مپŒهˆ©ç”¨هڈ¯èƒ½مپھ資و؛گم‚’ن½œم‚ٹه‡؛مپ™ç”ںو…‹ç³»م‚¨مƒ³م‚¸مƒ‹م‚¢مپ§مپ‚م‚‹مپ¨مپ„مپˆم‚‹[22]م€‚مپ“مپ®م‚ˆمپ†مپھç”ںو…‹ç³»مپ®ه¤‰هŒ–مپ¯م€په…±هکمپ™م‚‹ç¨®مپ¨ç’°ه¢ƒمپ®é€²هŒ–مپ«ه½±éں؟م‚’ن¸ژمپˆم‚‹[22]م€‚مپ“مپ®مپ“مپ¨مپ¯م€پوµ·مپ¨é™¸مپ®ه †ç©چ物مپ«و®‹مپ•م‚Œمپںç”ںç—•هŒ–çں³مپ‹م‚‰وکژم‚‰مپ‹مپ§مپ‚م‚‹م€‚مپمپ®ن»–مپ®ç”ں物و“¾ن¹±هٹ¹وœمپ«مپ¯م€په †ç©چ物مپ®مƒ†م‚¯م‚¹مƒپمƒ£مƒ¼مپ®ه¤‰هŒ– (ç¶ڑوˆگن½œç”¨)م€پç”ں物çپŒو¼‘(bioirrigation)م€پمپٹم‚ˆمپ³ه¾®ç”ں物مپ¨éç”ں物粒هگمپ®ç§»ه‹•مپھمپ©مپŒهگ«مپ¾م‚Œم‚‹م€‚ç”ں物و“¾ن¹±مپ¯ç”ں物çپŒو¼‘مپ®مƒ—مƒم‚»م‚¹مپ¨و··هگŒمپ•م‚Œم‚‹مپ“مپ¨مپŒمپ‚م‚‹مپŒم€پمپ“م‚Œم‚‰مپ®مƒ—مƒم‚»م‚¹مپ¯و··هگˆمپ™م‚‹م‚‚مپ®مپŒç•°مپھم‚‹م€‚ç”ں物çپŒو¼‘مپ¨مپ¯ه †ç©چ物ن¸مپ®و°´مپ¨و؛¶è³ھمپ®و··هگˆم‚’وŒ‡مپ—م€پç”ں物و“¾ن¹±مپ®م€Œهٹ¹وœم€چمپ«ç„¦ç‚¹م‚’ه½“مپ¦مپں用èھمپ§مپ‚م‚‹[20]م€‚ م‚»م‚¤م‚¦مƒپمپ¨م‚µم‚±مپ¯ç”ں物و“¾ن¹±م‚’و‹…مپ†ه¤§ه‹ç”ں物مپ®ن¾‹مپ§مپ‚م‚‹[23][24][25]م€‚مپ“م‚Œم‚‰مپ®ه¤§مپچمپھمƒم‚¯مƒمƒ•م‚،م‚¦مƒٹ(ه¤§ه‹ه؛•ç”ںç”ں物)مپ®ç”ں物و“¾ن¹±ç¨®مپ®و´»ه‹•مپ¯ç›®ç«‹مپ،م‚„مپ™مپ„مپŒم€په®ںç’°ه¢ƒمپ§م‚ˆم‚ٹه¤§مپچمپھه½±éں؟هٹ›م‚’وŒپمپ¤مپ®مپ¯م‚€مپ—م‚چم‚ˆم‚ٹه°ڈه‹مپ®ه¤ڑو¯›é،م€پم‚´مƒ¼م‚¹مƒˆم‚·مƒ¥مƒھمƒ³مƒ—م€پمƒمƒƒمƒ‰م‚·مƒ¥مƒھمƒ³مƒ—مپھمپ©مپ®ه°ڈمپ•مپھç„،è„ٹو¤ژه‹•ç‰©ï¼ˆمƒم‚¯مƒمƒ•م‚،م‚¦مƒٹ)مپ§مپ‚م‚‹[20][26]م€‚مپ“م‚Œم‚‰مپ®ه°ڈمپ•مپھç„،è„ٹو¤ژه‹•ç‰©مپ®و´»ه‹•مپ«مپ¯م€پç©´م‚’وژکمپ£مپںم‚ٹم€په †ç©چ物مپ®ç²’هگم‚’و‘‚هڈ–مپ—مپںم‚ٹم€پوژ’و³„مپ—مپںم‚ٹمپ™م‚‹مپ“مپ¨مپŒهگ«مپ¾م‚Œم€پو··هگˆمپ¨ه †ç©چ物مپ®و§‹é€ مپ®ه¤‰هŒ–مپ«ه¯„ن¸ژمپ™م‚‹م€‚ ç”ں物çپŒو¼‘ç”ں物çپŒو¼‘مپ¯ه…·ن½“çڑ„مپ«مپ¯م€په؛•ç”ںç”ں物مپŒه·£ç©´م‚’وژکم‚‹مپ“مپ¨مپ§ه †ç©چ物مپ®و·±éƒ¨مپ«وµ·و°´م‚’وµ¸é€ڈمپ•مپ›م‚‹مƒ—مƒم‚»م‚¹مپ§مپ‚م‚‹م€‚مپ“مپ®çµگوœمپ¨مپ—مپ¦ه †ç©چ物ن¸مپ®é–“éڑ™و°´مپ¨وµ·و°´مپ¨مپ®é–“مپ§و؛¶è§£ç‰©è³ھمپ®ن؛¤وڈ›مپŒèµ·مپ“م‚‹م€‚مپ“مپ®مƒ—مƒم‚»م‚¹مپ¯م€پوµ·و´‹مپ®ç”ں物هœ°çگƒهŒ–ه¦مپ®و–‡è„ˆمپ«مپٹمپ„مپ¦é‡چè¦پمپ§مپ‚م‚‹م€‚و²؟ه²¸مپ®و°´ç”ںç’°ه¢ƒمپ«مپ¯م€په †ç©چ物م‚’ن¸چه®‰ه®ڑمپ«مپ™م‚‹ç”ں物مپŒمپ„م‚‹مپ“مپ¨مپŒم‚ˆمپڈمپ‚م‚‹م€‚مپم‚Œم‚‰مپ¯ه †ç©چ物مپ®ç‰©çگ†çڑ„çٹ¶و…‹م‚’ه¤‰هŒ–مپ•مپ›م€پè‡ھè؛«م‚’هگ«م‚€ه¤ڑمپڈمپ®ç”ں物種مپ®ç”ںو´»ç’°ه¢ƒم‚’و•´مپˆو”¹ه–„مپ™م‚‹هٹ¹وœم‚’م‚‚مپںم‚‰مپ™م€‚مپ“م‚Œم‚‰مپ®ç”ں物مپ¯م€پç”ں物و“¾ن¹±م‚’ه¼•مپچèµ·مپ“مپ™مپ“مپ¨م‚‚ه¤ڑمپڈم€پم‚ˆمپڈç”ں物و“¾ن¹±مپ¨ç”ں物çپŒو¼‘مپ¨مپ„مپ†ç”¨èھمپ¯هگŒç¾©çڑ„مپ«ن½؟用مپ•م‚Œم‚‹مپ“مپ¨مپŒمپ‚م‚‹[27]م€‚ ç”ں物çپŒو¼‘مپ¯ م€Œç²’هگمپ®ه†چهٹ ه·¥م€چمپ¨م€Œوڈ›و°—م€چمپ¨مپ„مپ†2مپ¤مپ®ç•°مپھم‚‹مƒ—مƒم‚»م‚¹مپ¨مپ—مپ¦و©ں能مپ™م‚‹م€‚مپ“مپ®مƒ—مƒم‚»م‚¹مپ¯ه؛•ç”ںمپ®ه¤§ه‹ç„،è„ٹو¤ژه‹•ç‰©(é€ڑه¸¸مپ¯ç©´م‚’وژکم‚‹م‚‚مپ®) مپŒو‘‚é£ں (ه‹•ç‰©و€§و‘‚é£ں)م€پوژ’ن¾؟م€په·£ç©´مپ®و§‹ç¯‰م€پمپمپ—مپ¦ه‘¼هگ¸م‚’مپ™م‚‹مپ“مپ¨مپ§ه¼•مپچèµ·مپ“مپ•م‚Œم‚‹م€‚ç”ں物çپŒو¼‘مپ¯ه¤§é‡ڈمپ®é…¸ç´ مپ®è¼¸é€پ(酸هŒ–輸é€پ)م‚’و‹…مپ£مپ¦مپٹم‚ٹم€پç”ں物هœ°çگƒهŒ–ه¦çڑ„ه¾ھç’°مپ«ه¤§مپچمپھه½±éں؟م‚’ن¸ژمپˆمپ¦مپ„م‚‹م€‚ éپ و´‹و€§ه †ç©چ物
éپ و´‹و€§ه †ç©چ物(Pelagic sedimentsمپ¾مپںمپ¯pelagite)مپ¯م€پ陸هœ°مپ‹م‚‰éپ مپڈ離م‚Œمپںه¤–و´‹مپ®ه؛•مپ«ç²’هگمپŒو²ˆé™چمپ—مپںçµگوœمپ¨مپ—مپ¦è“„ç©چمپ™م‚‹ç´°ç²’مپ®ه †ç©چ物مپ§مپ‚م‚‹م€‚مپ“م‚Œم‚‰مپ®ç²’هگمپ¯م€پن¸»مپ«و¤چ物مƒ—مƒ©مƒ³م‚¯مƒˆمƒ³مپ¾مپںمپ¯ه‹•ç‰©مƒ—مƒ©مƒ³م‚¯مƒˆمƒ³مپ®ه¾®è¦–çڑ„مپھçں³çپ°è³ھمپ¾مپںمپ¯çڈھè³ھمپ®و®»مپ§م€پمپمپ—مپ¦ç²کهœںم‚µم‚¤م‚؛مپ®çڈھç •è³ھه †ç©چ物م€پمپ¾مپںمپ¯مپ“م‚Œم‚‰مپ®و··هگˆç‰©مپ‹م‚‰و§‹وˆگمپ•م‚Œمپ¦مپ„م‚‹م€‚ه¾®é‡ڈمپ®éڑ•çں³مپ®مپ،م‚ٹم‚„çپ«ه±±çپ°م‚‚م€پéپ و´‹ه †ç©چ物ه†…مپ§è¦‹م‚‰م‚Œم‚‹م€‚è»ںو³¥مپ®çµ„وˆگمپ«هں؛مپ¥مپ„مپ¦م€پéپ و´‹و€§ه †ç©چ物مپ«مپ¯ن¸»مپ« 3 مپ¤مپ®م‚؟م‚¤مƒ—مپŒهکهœ¨مپ—م€پمپم‚Œمپم‚Œçڈھè³ھè»ںو³¥م€پçں³çپ°è³ھè»ںو³¥م€پ赤色ç²کهœںمپ¨ه‘¼مپ°م‚Œم‚‹[29][30]م€‚ HMS مƒپمƒ£مƒ¬مƒ³م‚¸مƒ£مƒ¼هڈ· (1872–1876) مپ®èˆھوµ·ن»¥و¥م€پéپژهژ» 150ه¹´é–“مپ«م‚ڈمپںمپ£مپ¦م€پو·±وµ·مپ®مƒ—مƒم‚»م‚¹مپ¨ه †ç©چ物مپ«é–¢مپ™م‚‹ه؛ƒç¯„مپھç ”ç©¶مپŒè،Œم‚ڈم‚Œمپ¦مپچمپںم€‚مپ“مپ®èˆھوµ·مپ§مپ¯م€پوµ·ه؛•ه †ç©چ物مپ®وœ€هˆمپ®ن½“ç³»çڑ„مپھç ”ç©¶مپŒè،Œم‚ڈم‚Œمپں[31][32]م€‚مپمپ®ه…ˆé§†çڑ„مپھéپ ه¾پمپ‹م‚‰ن½•هچپه¹´م‚‚مپ®é–“م€پمپمپ—مپ¦ 20 ن¸–ç´€مپ®ه‰چهچٹم‚’é€ڑمپ—مپ¦م€پو·±وµ·مپ¯وœ¬è³ھçڑ„مپ«مپ¯éپ و´‹و€§مپ§مپ‚م‚‹مپ¨مپ؟مپھمپ›م‚‹مپ¨è€ƒمپˆم‚‰م‚Œمپ¦مپ„م‚‹م€‚ éپ و´‹ه †ç©چ物مپ®çµ„وˆگمپ¯م€پ3 مپ¤مپ®ن¸»مپھè¦په› مپ«م‚ˆمپ£مپ¦هˆ¶ه¾،مپ•م‚Œم‚‹م€‚1مپ¤ç›®مپ®è¦په› مپ¯م€پ陸ç”ںمپ¾مپںمپ¯هœںهœ°ç”±و¥مپ®ه †ç©چ物مپ«م‚ˆم‚‹ه¸Œé‡ˆمپ«ه½±éں؟مپ™م‚‹ن¸»è¦پمپھ陸هœ°مپ‹م‚‰مپ®è·é›¢مپ§مپ‚م‚‹م€‚ 2 مپ¤ç›®مپ®è¦په› مپ¯و°´و·±مپ§مپ‚م‚‹م€‚مپ“م‚Œمپ¯م€پم‚±م‚¤è³ھمپ¨çں³çپ°è³ھمپ®ن¸،و–¹مپ®ç”ںن½“ç²’هگمپŒوµ·ه؛•مپ«و²ˆé™چمپ™م‚‹éڑ›مپ®ن؟هکمپ«ه½±éں؟م‚’ن¸ژمپˆم‚‹م€‚3مپ¤ç›®مپ®è¦په› مپ¯وµ·مپ®è‚¥و²ƒه؛¦ï¼ˆç”ں物ç”ں産و€§م€پمƒگم‚¤م‚ھمƒم‚¹ï¼‰مپ§مپ‚م‚ٹم€پè،¨ه±¤و°´مپ§ç”ںوˆگمپ•م‚Œم‚‹ç”ں物起و؛گمپ®ç²’هگمپ®é‡ڈم‚’هˆ¶ه¾،مپ™م‚‹[29][30]م€‚ م‚؟مƒ¼مƒ“مƒ€م‚¤مƒˆ
م‚؟مƒ¼مƒ“مƒ€م‚¤مƒˆمپ¯و؟پوµپمپ®هœ°è³ھه †ç©چ物مپ§مپ‚م‚ٹم€پوµپن½“مپ¨ه †ç©چ物é‡چهٹ›وµپمپ®èچهگˆمپ®ن¸€ç¨®مپ§مپ‚م‚ٹم€پ膨ه¤§مپھé‡ڈمپ®ç •ه±‘ه †ç©چ物م‚’و·±وµ·مپ¸مپ¨é€پم‚ٹè¾¼م‚€م€‚م‚؟مƒ¼مƒ“مƒ€م‚¤مƒˆمپ¯م€په¤§é™¸و£ڑمپ®ç«¯مپ®و€¥مپھو–œé¢م‚’و»‘م‚ٹèگ½مپ،م‚‹و°´ن¸مپ®é›ھه´©مپ«م‚ˆمپ£مپ¦م€په¤§é™¸و£ڑمپ®ن¸‹مپ®و·±وµ·مپ®è°·م€پمپ¾مپںمپ¯و·±مپ„و¹–مپ®هگŒو§کمپ®و§‹é€ مپ«ه †ç©چمپ™م‚‹م€‚物è³ھمپŒوµ·مپ®è°·مپ«و²ˆم‚€مپ¨مپچم€پوœ€هˆمپ«و²ˆé™چمپ™م‚‹مپ®مپ¯ç ‚م‚„مپمپ®ن»–مپ®ç²—مپ„物è³ھمپ§مپ‚م‚ٹم€پو¬،مپ«و³¥م€پمپمپ—مپ¦وœ€çµ‚çڑ„مپ«مپ¯éه¸¸مپ«ç´°مپ‹مپ„ç²’هگçٹ¶ç‰©è³ھمپŒو²ˆé™چمپ™م‚‹م€‚مپ“مپ®ه †ç©چمپ®é †ç•ھمپ¯م€پمپ“م‚Œم‚‰مپ®ه²©çں³م‚’特ه¾´ن»کمپ‘م‚‹مƒœم‚¦مƒ م‚·مƒ¼م‚±مƒ³م‚¹ï¼ˆBouma sequences)م‚’ن½œوˆگمپ™م‚‹م€‚ م‚؟مƒ¼مƒ“مƒ€م‚¤مƒˆمپ¯ 1950ه¹´ن»£مپ«هˆم‚پمپ¦èھچèکمپ•م‚Œ[34]م€پوœ€هˆمپ®ç›¸مƒ¢مƒ‡مƒ«مپ¯ 1962ه¹´مپ«Boumaمپ«م‚ˆمپ£مپ¦é–‹ç™؛مپ•م‚Œمپں[35]م€‚مپمپ®و™‚ن»¥و¥م€پم‚؟مƒ¼مƒ“مƒ€م‚¤مƒˆمپ¯وœ€م‚‚集ن¸çڑ„مپ«ç ”究مپ•م‚Œمپ¦مپ„م‚‹و·±وµ·ه †ç©چ物相مپ®1مپ¤مپ§مپ‚م‚ٹم€پو·±وµ·م‚„هœ°ن¸‹ï¼ˆç‚هŒ–و°´ç´ )مƒœم‚¢مƒ›مƒ¼مƒ«م€پ陸ن¸ٹمپ«éœ²ه‡؛مپ—مپ¦مپ„م‚‹هڈ¤ن»£مپ®éœ²é مپھمپ©مپ®ه †ç©چ物م‚³م‚¢مپ‹م‚‰م‚ˆمپڈçں¥م‚‰م‚Œمپ¦مپ„م‚‹م€‚特ه®ڑمپ®م‚؟مƒ¼مƒ“مƒ€م‚¤مƒˆç³»م‚’èھ؟مپ¹م‚‹مپ“مپ¨مپ«م‚ˆم‚ٹم€پمپمپ®هœ°ç‚¹مپ«مپٹمپ‘م‚‹ه †ç©چ物特و€§م‚„ه±¤ç›¸م‚’وکژم‚‰مپ‹مپ«مپ™م‚‹مپ“مپ¨مپŒمپ§مپچم‚‹[36][37]م€‚ م‚³مƒ³م‚؟مƒ¼مƒ©م‚¤مƒˆï¼ˆç‰و·±ه؛¦وµپه †ç©چ物)
م‚³مƒ³م‚؟مƒ¼مƒ©م‚¤مƒˆï¼ˆcontourite)مپ¯م€پن¸€èˆ¬çڑ„مپ«ه¤§é™¸éڑ†èµ·مپ‹م‚‰ن½ژو–œé¢مپ«ه½¢وˆگمپ•م‚Œم‚‹ه †ç©چ物مپ§مپ‚م‚‹مپŒم€پهµگمپ®و³¢مپ®هں؛部م‚ˆم‚ٹن¸‹مپ«مپ‚م‚‹ه ´و‰€مپھم‚‰مپ©مپ“مپ§م‚‚ç™؛ç”ںمپ™م‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹م€‚م‚³مƒ³م‚؟مƒ¼مƒ©م‚¤مƒˆمپ¯ç†±ه،©èھکèµ·مپ®و·±وµ·ه؛•وµپمپ«م‚ˆمپ£مپ¦ç”ںوˆگمپ•م‚Œم€پ風م‚„و½®و±گهٹ›مپ®ه½±éں؟م‚’هڈ—مپ‘م‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹[39][40]م€‚م‚³مƒ³م‚؟مƒ¼مƒ©م‚¤مƒˆه †ç©چ物مپ®هœ°ه½¢مپ¯م€پن¸»مپ«و·±وµ·ه؛•وµپé€ںه؛¦م€په †ç©چ物ن¾›çµ¦م€پمپٹم‚ˆمپ³وµ·ه؛•هœ°ه½¢مپ®ه½±éں؟م‚’هڈ—مپ‘م‚‹[41]م€‚ م‚³مƒ³م‚؟مƒ¼مƒ©م‚¤مƒˆمپ¯م€پ1960ه¹´ن»£هˆé مپ«مƒ–مƒ«مƒ¼م‚¹مƒ»مƒ’مƒ¼م‚¼مƒ³مپ¨م‚¦مƒƒم‚؛مƒ›مƒ¼مƒ«وµ·و´‹ç ”究و‰€مپ®هگŒهƒڑمپ«م‚ˆمپ£مپ¦وœ€هˆمپ«ç‰¹ه®ڑمپ•م‚Œمپںم€‚ه½¼م‚‰مپ®é‡چè¦پمپھè«–و–‡[42]مپ¯م€پهŒ—م‚¢مƒ،مƒھم‚«و±éƒ¨و²–مپ®ه¤§é™¸éڑ†èµ·مپ®ه½¢وˆگمپ«مپٹمپ„مپ¦م€پç‰é«کç·ڑمپ«و²؟مپ£مپںوµ·ه؛•وµپمپŒéه¸¸مپ«é‡چè¦پمپھه½±éں؟م‚’هڈٹمپ¼مپ—مپ¦مپ„م‚‹مپ“مپ¨م‚’ç¤؛مپ—مپںم€‚مپ“م‚Œم‚‰مپ®هچٹو°¸ن¹…çڑ„مپھو–œé¢مپ«و²؟مپ£مپںوµپم‚Œمپ®ه †ç©چ物مپ¯م€پمپ™مپگمپ«م‚³مƒ³م‚؟مƒ¼مƒ©م‚¤مƒˆمپ¨مپ—مپ¦çں¥م‚‰م‚Œم‚‹م‚ˆمپ†مپ«مپھم‚ٹم€پç‰é«کç·ڑمپ§ن¸»مپ«و§‹وˆگمپ•م‚Œمپںو–œé¢مپ«ه¹³è،Œمپ§ç´°é•·مپڈç››م‚ٹن¸ٹمپŒمپ£مپںه †ç©چن½“مپ®ه¢ƒç•Œمپ¯م€پç‰é«کç·ڑمƒ‰مƒھمƒ•مƒˆمپ¨مپ—مپ¦çں¥م‚‰م‚Œم‚‹م‚ˆمپ†مپ«مپھمپ£مپں[43][44]م€‚ هچٹéپ و´‹و€§ه †ç©چ物هچٹéپ و´‹و€§ه †ç©چ物(Hemipelagic sedimentsم€پhemipelagite)مپ¯م€په †ç©چ物مپ«وœ€م‚‚è؟‘مپ„陸ه،ٹمپ¾مپںمپ¯و°´ن¸مپ«ç”ںوپ¯مپ™م‚‹ç”ں物مپ«ç”±و¥مپ™م‚‹é™¸ç”ںمپٹم‚ˆمپ³مپ„مپڈمپ¤مپ‹مپ®ç”ں物起و؛گ物è³ھمپ§مپ‚م‚‹ç²کهœںمپٹم‚ˆمپ³م‚·مƒ«مƒˆم‚µم‚¤م‚؛مپ®ç²’هگمپ§و§‹وˆگمپ•م‚Œم‚‹وµ·و´‹ه †ç©چ物مپ®ن¸€ç¨®مپ§مپ‚م‚‹[45][46]م€‚هچٹéپ و´‹و€§ه †ç©چ物مپ¯ه¤§é™¸و£ڑمپ¨ه¤§é™¸éڑ†èµ·مپ«ه †ç©چمپ—م€پéپ و´‹و€§ه †ç©چ物مپ¨مپ¯çµ„وˆگمپŒç•°مپھم‚‹م€‚éپ و´‹ه †ç©چ物مپ¯م€پن¸»مپ«و°´وں±مپ¾مپںمپ¯وµ·ه؛•مپ«ç”ںوپ¯مپ™م‚‹ç”ں物مپ‹م‚‰مپ®ç”ں物起و؛گمپ®ç‰©è³ھمپ§و§‹وˆگمپ•م‚Œمپ¦مپٹم‚ٹم€پ陸ç”ں物è³ھمپ¯مپ»مپ¨م‚“مپ©مپ¾مپ£مپںمپڈهگ«مپ¾م‚Œمپ¦مپ„مپھمپ„[45]م€‚陸ç”ں物è³ھمپ«مپ¯م€پé•·çں³م‚„çں³è‹±مپھمپ©مپ®مƒھم‚½م‚¹مƒ•م‚§م‚¢مپ‹م‚‰مپ®é‰±ç‰©مپŒهگ«مپ¾م‚Œم‚‹م€‚陸ن¸ٹمپ®çپ«ه±±و´»ه‹•م€پ風مپ«هگ¹مپ‹م‚Œمپںه †ç©چ物م€پمپٹم‚ˆمپ³ه·مپ‹م‚‰و”¾ه‡؛مپ•م‚Œمپںç²’هگçٹ¶ç‰©è³ھمپ¯م€پهچٹéپ و´‹ه †ç©چ物مپ«ه¯„ن¸ژمپ™م‚‹هڈ¯èƒ½و€§مپŒمپ‚م‚‹[47]م€‚مپ“م‚Œم‚‰مپ®ه †ç©چ物مپ¯م€پو°—ه€™ه¤‰ه‹•م‚’特ه®ڑمپ—م€په †ç©چ物起و؛گمپ®ه¤‰هŒ–م‚’特ه®ڑمپ™م‚‹مپںم‚پمپ«هˆ©ç”¨مپ§مپچم‚‹[48][49]م€‚
ç”ںو…‹ه¦مƒ™مƒ³مƒˆم‚¹(خ²خخ½خ¸خ؟د‚ (bأ©nthos)م€پهڈ¤ن»£م‚®مƒھم‚·مƒ£èھمپ§م€Œthe depths (of the sea)م€چم‚’و„ڈه‘³مپ™م‚‹ )مپ¯م€پوµ·ه؛•ن¸ٹ(ه؛•ç”ںه¸¯ï¼‰م‚„وµ·ه؛•ن¸م€پمپ¾مپںمپ¯مپمپ®è؟‘مپڈمپ«ç”ںوپ¯مپ™م‚‹ç”ں物مپ®ç¾¤é›†مپ§مپ‚م‚‹م€‚特مپ«ه †ç©چ物مپ®مپ™مپگن¸ٹمپ§ç”ںوپ¯مپ™م‚‹ç”ں物مپ¯Hyperbenthos(hupأ©rمپ¯هڈ¤ن»£م‚®مƒھم‚·مƒ£èھمپ§'over'م‚’و„ڈه‘³مپ™م‚‹ï¼‰م€په †ç©چ物مپ®ن¸ٹ部مپ«ç”ںوپ¯مپ™م‚‹ç”ں物مپ¯Epibenthos(epأ مپ¯â€™on top of’مپ®و„ڈه‘³ï¼‰م€په †ç©چ物مپ«ç©´م‚’ه½«م‚ٹه†…部مپ«ç”ںوپ¯مپ™م‚‹ç”ں物مپ¯Endobenthos(أ©ndonمپ¯'inner'مپ®و„ڈه‘³ï¼‰مپ¨ه‘¼مپ°م‚Œم‚‹م€‚ مƒںم‚¯مƒمƒ™مƒ³مƒˆم‚¹
وµ·و´‹مƒںم‚¯مƒمƒ™مƒ³مƒˆم‚¹مپ¯م€پوµ·و´‹مپ®ه؛•ç”ںه¸¯مپ«ç”ںوپ¯مپ™م‚‹ه¾®ç”ں物مپ§مپ‚م‚ٹم€پوµ·ه؛•مپ®è؟‘مپڈم‚„è،¨ه±¤ه †ç©چ物ه†…مپ«ç”ںوپ¯مپ—مپ¦مپ„م‚‹م€‚مƒ™مƒ³مƒˆم‚¹مپ¨مپ„مپ†è¨€è‘‰مپ¯م‚®مƒھم‚·مƒ£èھمپ«ç”±و¥مپ—م€پم€Œوµ·مپ®و·±مپ„و‰€م€چم‚’و„ڈه‘³مپ™م‚‹م€‚ه؛•ç”ںه¾®ç”ں物مپ¯ه¤§é™¸و£ڑمپ®وµ·ه؛•ن¸ٹم‚„مپمپ®ه‘¨è¾؛مپ§مپ¯مپ„مپںم‚‹مپ¨مپ“م‚چمپ«è¦‹م‚‰م‚Œم€پو·±وµ·مپ مپ‘مپ§مپھمپڈم€پوµ·ه؛•ه †ç©چ物ه†…م‚„ه †ç©چ物ن¸ٹمپ§م‚ˆم‚ٹه¤ڑو§کمپ§مپ‚م‚‹م€‚وµ…瀬مپ§مپ¯م€پوµ·èچ‰مپ®ç¹پ茂هœ°م€پم‚µمƒ³م‚´ç¤پم€پم‚±مƒ«مƒ—مپ®و£®مپŒç‰¹مپ«è±ٹمپ‹مپھç”ںوپ¯هœ°م‚’وڈگن¾›مپ™م‚‹م€‚وœ‰ه…‰ه¸¯مپ§مپ¯م€په؛•ç”ںçڈھè—»مپŒه…‰هگˆوˆگç”ں物مپ¨مپ—مپ¦ه„ھه‹¢مپ§مپ‚م‚‹م€‚و½®é–“ه¸¯مپ§مپ¯م€پو½®مپ®ه¤‰هŒ–مپŒمƒم‚¤م‚¯مƒمƒ™مƒ³مƒˆم‚¹مپ®è‚²وˆگمپ«ه¤§مپچمپھه½±éں؟م‚’ن¸ژمپˆم‚‹م€‚
çڈھè—»مپ¯م€پن¸»مپ«هچکç´°èƒè—»é،مپ‹م‚‰مپھم‚‹ç´„100,000種مپŒçں¥م‚‰م‚Œمپ¦مپٹم‚ٹم€پé–€م‚’ه½¢وˆگمپ™م‚‹(مپںمپ مپ—م€پمپ“مپ®و•°مپ«مپ¯è°è«–مپŒمپ‚م‚‹)م€‚çڈھè—»مپ¯م€پهœ°çگƒن¸ٹمپ§و¯ژه¹´ç”ںوˆگمپ•م‚Œم‚‹é…¸ç´ مپ®ç´„20% م‚’ç”ںوˆگمپ—[52]م€پçڈھè—»مپŒç”ںوپ¯مپ™م‚‹و°´هںںمپ‹م‚‰و¯ژه¹´ 67ه„„مƒˆمƒ³ن»¥ن¸ٹمپ®م‚±م‚¤ç´ م‚’هڈ–م‚ٹè¾¼مپ؟[53]م€پوµ·و´‹مپ«è¦‹م‚‰م‚Œم‚‹وœ‰و©ں物è³ھمپ®مپ»مپ¼هچٹهˆ†م‚’هچ م‚پمپ¦مپ„م‚‹م€‚  ه††çں³è—»مپ¯م€پ移ه‹•ç”¨مپ® 2 مپ¤مپ®éو¯›م‚’وŒپمپ¤ه¾®ç´°مپھهچکç´°èƒمپ®ه…‰هگˆوˆگهژںç”ںç”ں物مپ§مپ‚م‚‹م€‚مپم‚Œم‚‰مپ®مپ»مپ¨م‚“مپ©مپ¯م€پcoccolithsمپ¨ه‘¼مپ°م‚Œم‚‹èڈ¯م‚„مپ‹مپھه††ه½¢مپ®مƒ—مƒ¬مƒ¼مƒˆمپ¾مپںمپ¯م‚¹م‚±مƒ¼مƒ«مپ§è¦†م‚ڈم‚Œمپںو®»مپ«م‚ˆمپ£مپ¦ن؟è·مپ•م‚Œمپ¦مپ„م‚‹م€‚Coccolithsمپ¯ç‚é…¸م‚«مƒ«م‚·م‚¦مƒ مپ‹م‚‰مپ§مپچمپ¦مپ„م‚‹م€‚ه††çں³è—»مپ®è‹±èھهگچمپ§مپ‚م‚‹coccolithophore مپ¨مپ„مپ†ç”¨èھمپ¯م€پم€Œç¨®هگم‚’éپ‹مپ¶çں³م€چم‚’و„ڈه‘³مپ™م‚‹م‚®مƒھم‚·مƒ£èھمپ«ç”±و¥مپ—م€پمپمپ®ه°ڈمپ•مپھم‚µم‚¤م‚؛مپ¨مپم‚Œم‚‰مپŒéپ‹مپ¶م‚³م‚³مƒھم‚¹çں³مپ®مپ“مپ¨م‚’وŒ‡مپ—مپ¦مپ„م‚‹م€‚éپ©هˆ‡مپھو،ن»¶ن¸‹مپ§مپ¯م€پن»–مپ®و¤چ物مƒ—مƒ©مƒ³م‚¯مƒˆمƒ³مپ¨هگŒو§کمپ«مƒ–مƒ«مƒ¼مƒ م‚’ه½¢وˆگمپ—م€پوµ·م‚’ن¹³ç™½è‰²مپ«ه¤‰مپˆم‚‹مپ“مپ¨مپŒمپ§مپچم‚‹[54]م€‚
و”¾و•£è™«مپ¯هچکç´°èƒمپ®وچ•é£ںو€§هژںç”ںç”ں物مپ§مپ‚م‚‹م€‚é€ڑه¸¸مپ¯م‚·مƒھم‚«مپ§مپ§مپچمپںم€په¾®ه°ڈمپھç©´مپŒé–‹مپ„مپںç²¾ه·§مپھçگƒçٹ¶مپ®و®»مپ«هŒ…مپ¾م‚Œمپ¦مپ„م‚‹م€‚英èھمپ®هگچه‰چ(Radiolarians)مپ¯مƒ©مƒ†مƒ³èھمپ®م€Œهچٹه¾„م€چ(radius)مپ«ç”±و¥مپ™م‚‹م€‚ç©´مپ‹م‚‰ن½“مپ®ن¸€éƒ¨م‚’ن¼¸مپ°مپ—مپ¦çچ²ç‰©م‚’وچ•م‚‰مپˆم‚‹م€‚çڈھè—»مپ®م‚·مƒھم‚«مƒ•مƒ©م‚¹مƒˆمƒ«مپ¨هگŒو§کمپ«م€پو”¾و•£è™«مپŒو»م‚“مپ§وµ·و´‹ه †ç©چ物مپ®ن¸€éƒ¨مپ¨مپ—مپ¦ن؟هکمپ•م‚Œم‚‹مپ¨م€پو”¾و•£è™«مپ®و®»مپ¯وµ·ه؛•مپ«و²ˆم‚€م€‚مپ“م‚Œم‚‰مپ®هŒ–çں³مپ¯ه¾®هŒ–çں³مپ¨مپ—مپ¦م€پéپژهژ»مپ®وµ·و´‹çٹ¶و³پمپ«é–¢مپ™م‚‹è²´é‡چمپھوƒ…ه ±م‚’وڈگن¾›مپ™م‚‹[55]م€‚
و”¾و•£è™«مپ®م‚ˆمپ†مپ«م€پوœ‰ه”虫مپ¯هچکç´°èƒمپ®وچ•é£ںو€§هژںç”ںç”ں物مپ§مپ‚م‚ٹم€پç©´مپ®مپ‚م‚‹و®»مپ§ن؟è·مپ•م‚Œمپ¦مپ„م‚‹م€‚ه½¼م‚‰مپ®هگچه‰چforaminiferansمپ¯مƒ©مƒ†مƒ³èھمپ®م€Œç©´م‚’وژکم‚‹ن؛؛م€چ(hole bearers)مپ«ç”±و¥مپ™م‚‹م€‚مپ—مپ°مپ—مپ°مƒ†م‚¹مƒˆمپ¨ه‘¼مپ°م‚Œم‚‹ه½¼م‚‰مپ®و®»مپ«مپ¯مƒپمƒ£مƒ³مƒگمƒ¼مپŒمپ‚م‚‹ï¼ˆوœ‰ه”虫مپ¯وˆگé•·مپ™م‚‹مپ«مپ¤م‚Œمپ¦مƒپمƒ£مƒ³مƒگمƒ¼م‚’è؟½هٹ مپ—مپ¦مپ„مپڈ)م€‚و®»مپ¯é€ڑه¸¸و–¹è§£çں³مپ§مپ§مپچمپ¦مپ„م‚‹مپŒم€په‡é›†مپ—مپںه †ç©چ物粒هگمپ¾مپںمپ¯م‚مƒˆمƒ³م€پمپٹم‚ˆمپ³ï¼ˆمپ¾م‚Œمپ«ï¼‰م‚·مƒھم‚«مپ§مپ§مپچمپ¦مپ„م‚‹ه ´هگˆم‚‚مپ‚م‚‹م€‚مپ»مپ¨م‚“مپ©مپ®وœ‰ه”虫مپ¯ه؛•ç”ںو€§مپ§مپ‚م‚‹مپŒم€پç´„40種مپ¯وµ®éپٹو€§مپ§مپ‚م‚‹[56]م€‚مپم‚Œم‚‰مپ¯م€پ科ه¦è€…مپŒéپژهژ»مپ®ç’°ه¢ƒم‚„و°—ه€™مپ«مپ¤مپ„مپ¦ه¤ڑمپڈمپ®مپ“مپ¨م‚’وژ¨و¸¬مپ§مپچم‚‹م‚ˆمپ†مپ«مپ™م‚‹م€پهچپهˆ†مپ«ç¢؛ç«‹مپ•م‚ŒمپںهŒ–çں³è¨ک録مپ§ه؛ƒمپڈç ”ç©¶مپ•م‚Œمپ¦مپ„م‚‹[55]م€‚ وœ‰ه”虫مپ¨çڈھè—»مپ¯مپ©مپ،م‚‰م‚‚وµ®éپٹو€§مپ¨ه؛•ç”ںو€§مپ®ه½¢و…‹م‚’وŒپمپ£مپ¦مپ„م‚‹م€‚مپ¤مپ¾م‚ٹم€پو°´وں±مپ«و¼‚مپ£مپںم‚ٹم€پوµ·ه؛•مپ®ه †ç©چ物مپ«ç”ںوپ¯مپ—مپںم‚ٹمپ§مپچم‚‹م€‚مپ„مپڑم‚Œمپ«مپ›م‚ˆم€پو»م‚“مپ ه¾Œم€پè²و®»مپ¯وµ·ه؛•مپ«èگ½مپ،مپ¦م‚†مپڈم€‚مپ“م‚Œم‚‰مپ®م‚·م‚§مƒ«مپ¯م€پو°—ه€™مپ®مƒ—مƒم‚م‚·مپ¨مپ—مپ¦ه؛ƒمپڈن½؟用مپ•م‚Œمپ¦مپ„م‚‹م€‚è²و®»مپ®هŒ–ه¦çµ„وˆگمپ¯م€پè²و®»مپŒه½¢وˆگمپ•م‚Œمپںمپ¨مپچمپ®وµ·و´‹مپ®هŒ–ه¦çµ„وˆگمپ®çµگوœمپ§مپ‚م‚‹م€‚éپژهژ»مپ®و°´و¸©مپ¯م€پم‚·م‚§مƒ«ه†…مپ®ه®‰ه®ڑمپ—مپںé…¸ç´ هگŒن½چن½“مپ®و¯”çژ‡مپ‹م‚‰وژ¨و¸¬مپ™م‚‹مپ“مپ¨م‚‚مپ§مپچم‚‹م€‚مپ“م‚Œمپ¯م€پ軽مپ„هگŒن½چن½“مپŒو¸©مپ‹مپ„و°´ن¸مپ§م‚ˆم‚ٹه®¹وک“مپ«è’¸ç™؛مپ—م€پé‡چمپ„هگŒن½چن½“مپŒم‚·م‚§مƒ«ه†…مپ«و®‹م‚‹مپںم‚پمپ§مپ‚م‚‹م€‚éپژهژ»مپ®و°—ه€™مپ«é–¢مپ™م‚‹وƒ…ه ±مپ¯م€پوœ‰ه”虫مپ¨çڈھè—»مپ®è±ٹه¯Œمپ•مپ‹م‚‰مپ•م‚‰مپ«وژ¨و¸¬مپ§مپچم‚‹[57]م€‚ 6600 ن¸‡ه¹´ه‰چمپ«وپگç«œم‚’絶و»…مپ•مپ›مپںçھپ然مپ®çµ¶و»…مپ¯م€پن»–مپ®مپ™مپ¹مپ¦مپ®ه‹•و¤چ物種مپ® 4 هˆ†مپ® 3 م‚’絶و»…مپ•مپ›مپںم€‚مپ—مپ‹مپ—م€پمپمپ®ه¾Œم€پو·±وµ·مپ®ه؛•ç”ںوœ‰ه”虫مپŒç¹پو „مپ—مپںم€‚ 2020ه¹´مپ«مپ¯م€پç ”ç©¶è€…مپŒمپ“م‚Œم‚‰مپ®ه؛•ç”ںوœ‰ه”虫مپ®ن½•هچƒم‚‚مپ®م‚µمƒ³مƒ—مƒ«مپ®هŒ–ه¦çµ„وˆگم‚’èھ؟وں»مپ—م€پمپمپ®çµگوœم‚’ن½؟用مپ—مپ¦مپ“م‚Œمپ¾مپ§مپ§وœ€م‚‚詳細مپھهœ°çگƒمپ®و°—ه€™è¨ک録م‚’ن½œوˆگمپ—مپںمپ“مپ¨مپŒه ±ه‘ٹمپ•م‚Œمپں[58][59]م€‚ مپ„مپڈمپ¤مپ‹مپ®ه²©çں³ه†…ه¾®ç”ں物(endoliths)مپ¯éه¸¸مپ«é•·مپ„ه¯؟ه‘½م‚’وŒپمپ£مپ¦مپ„م‚‹م€‚ 2013ه¹´مپ«م€پç ”ç©¶è€…مپ¯وµ·ه؛•مپ«مپٹمپم‚‰مپڈو•°ç™¾ن¸‡ه¹´ه‰چمپ®endolithsمپ®è¨¼و‹ م‚’ه ±ه‘ٹمپ—م€پç”ںوˆگو™‚é–“مپ¯10,000ه¹´مپ§مپ‚مپ£مپں[60]م€‚مپ“م‚Œم‚‰مپ¯م‚†مپ£مپڈم‚ٹمپ¨ن»£è¬مپ•م‚Œم€پن¼‘çœ çٹ¶و…‹مپ§مپ¯مپھمپ„م€‚م‚·مƒ™مƒھم‚¢مپ§è¦‹مپ¤مپ‹مپ£مپںمپ„مپڈمپ¤مپ‹مپ®و”¾ç·ڑèڈŒمپ¯م€پ50 ن¸‡ه¹´ه‰چمپ®م‚‚مپ®مپ¨وژ¨ه®ڑمپ•م‚Œمپ¦مپ„م‚‹[61][62][63]م€‚ ه †ç©چ物م‚³م‚¢ هڈ³مپ®ه›³مپ¯م€په †ç©چ物م‚³م‚¢مپ®ن¾‹م‚’ç¤؛مپ—مپ¦مپ„م‚‹م€‚مپ“مپ®م‚µمƒ³مƒ—مƒ«مپ¯م€پ2018ه¹´é ƒمپ«م‚¦مƒڑمƒ«مƒٹمƒ“م‚¯مƒ•م‚£مƒ¨مƒ«مƒ‰ï¼ˆUpernavik Fjord)مپ‹م‚‰ه›هڈژمپ•م‚Œمپںم€‚ç²’ه؛¦و¸¬ه®ڑمپŒè،Œم‚ڈم‚Œم€پن¸ٹ部 50 cm مپŒ210Pbو³•مپ«م‚ˆم‚ٹه¹´ن»£و¸¬ه®ڑمپ•م‚Œمپں[64]م€‚  ç‚ç´ ه¾ھç’°  وµ·و´‹ç‚ç´ مپ¨ç‚ç´ éڑ”離مپ«مپ¤مپ„مپ¦مپ®è€ƒمپˆو–¹مپ¯م€پè؟‘ه¹´م€پو§‹é€ مپ«هں؛مپ¥مپڈهŒ–ه¦هڈچه؟œو€§مپ®è¦³ç‚¹مپ‹م‚‰م€پوœ‰و©ںç‚ç´ مپ®هˆ†è§£é€ںه؛¦مپ«مپٹمپ‘م‚‹ç”ںو…‹ç³»مپ®ه½¹ه‰²م‚’هگ«م‚€è¦³ç‚¹مپ«ç§»è،Œمپ—مپ¦مپ„م‚‹[68]م€‚وœ‰و©ںç‚ç´ مپ¨ç”ںو…‹ç³»مپ®é–¢ن¸ژمپ«é–¢مپ™م‚‹مپ“مپ®è¦‹و–¹مپ®ه¤‰هŒ–مپ¯م€پç”ں物ه¦مپ«مپٹمپ‘م‚‹هˆ†هگç”ں物ه¦çڑ„مپھم‚¢مƒ—مƒمƒ¼مƒپمپ®ه¼·هŒ–م‚„م€پç”ںه‘½مپ®é™گç•Œمپ«é–¢مپ™م‚‹ç™؛見م€په®ڑé‡ڈçڑ„مƒ¢مƒ‡مƒھمƒ³م‚°مپ®é€²و©م€پوµ·و´‹ç‚ç´ ه¾ھç’°مپ®هڈ¤ç ”究م€پو–°مپ—مپ„هˆ†وگوٹ€è،“م€پمپمپ—مپ¦مپم‚Œم‚‰مپ«ن؟‚م‚ڈم‚‹و§کم€…مپھه¦éڑ›çڑ„مپھهڈ–م‚ٹ組مپ؟مپ«م‚ˆم‚‹مپ¨مپ“م‚چمپŒه¤§مپچمپ„م€‚2020ه¹´م€پLaRoweم‚‰مپ¯وµ·و´‹ه †ç©چ物مپ¨هœ°çگƒè¦ڈو¨،مپ®ç‚ç´ ه¾ھç’°مپ«é–¢é€£مپ™م‚‹م€پ複و•°مپ®ç§‘ه¦هˆ†é‡ژمپ«مپ¾مپںمپŒم‚‹مپ“مپ®ه•ڈé،Œمپ®ه؛ƒمپ„視é‡ژم‚’و¦‚èھ¬مپ—مپ¦مپ„م‚‹[69][68]م€‚ هœ°çگƒهڈ²هژںهˆمپ®هœ°çگƒمپ§مپ¯مپ¾مپڑم€پو¥µç«¯مپھçپ«ه±±و´»ه‹•مپ¨ن»–مپ®ه¤©ن½“مپ¨مپ®é »ç¹پمپھè،çھپمپ«م‚ˆم‚ٹم€پهœ°çگƒمپ¯و؛¶مپ‘مپ¦مپ„مپںم€‚مپ—مپ مپ„مپ«وƒ‘وکںمپ®ه¤–ه±¤مپŒه†·مپˆمپ¦ه›؛مپ„هœ°و®»م‚’ه½¢وˆگمپ—م€په¤§و°—ن¸مپ«و°´مپŒè“„ç©چمپ—ه§‹م‚پمپںم€‚م‚¬م‚¹و”¾ه‡؛مپ¨çپ«ه±±و´»ه‹•مپŒهژںه§‹ه¤§و°—م‚’ن½œم‚ٹه‡؛مپ—مپںم€‚ه½—وکںمپ‹م‚‰éپ‹مپ°م‚Œمپںو°·مپ«م‚ˆمپ£مپ¦ه¼·هŒ–مپ•م‚Œمپںه‡ç¸®و°´è’¸و°—مپ¯م€پوµ·م‚’ن½œم‚ٹه‡؛مپ—مپں[70][71][72]م€‚ ç´„40ه„„ه¹´ه‰چمپ®ه§‹ç”ںن»£مپ®é–‹ه§‹مپ¾مپ§مپ«م€په²©çں³مپ¯مپ—مپ°مپ—مپ°ç،¬ç ‚ه²©م€پو³¥ه²©م€پçپ«ه±±و€§ه †ç©چ物م€په¸¯çٹ¶é‰„ه±¤مپھمپ©مپ®é«که؛¦مپ«ه¤‰وˆگمپ—مپںو·±وµ·ه †ç©چ物مپ§مپ‚مپ£مپںم€‚م‚°مƒھمƒ¼مƒ³م‚¹مƒˆمƒ¼مƒ³ مƒ™مƒ«مƒˆمپ¯ه…¸ه‹çڑ„مپھه¤ھهڈ¤مپ®هœ°ه±¤مپ§مپ‚م‚ٹم€پé«که“پن½چمپ¨ن½ژه“پن½چمپ®ه¤‰وˆگه²©مپŒن؛¤ن؛’مپ«ن¸¦م‚“مپ§مپ„م‚‹م€‚é«که“پن½چمپ®ه²©مپ¯çپ«ه±±ه³¶مپ®ه¼§مپ«ç”±و¥مپ—م€پن½ژه“پن½چمپ®ه¤‰وˆگه²©مپ¯م€پéڑ£وژ¥مپ™م‚‹ه³¶مپ®ه²©مپ‹م‚‰وµ¸é£ںمپ•م‚Œم€په‰چه¼§ç›†هœ°مپ«ه †ç©چمپ—مپںو·±وµ·ه †ç©چ物م‚’è،¨مپ—مپ¦مپ„م‚‹[73]م€‚çں¥م‚‰م‚Œمپ¦مپ„م‚‹وœ€هڈ¤مپ®è¶…ه¤§é™¸مƒمƒ‡م‚£مƒ‹م‚¢مپ¯م€پç´„10ه„„ه¹´ه‰چمپ«ه½¢وˆگمپ•م‚Œم€پç´„2ه„„5000ن¸‡ه¹´ه¾Œمپ®هژںç”ںن»£مپ®ه¾Œهچٹمپ«هˆ†è£‚مپ—ه§‹م‚پمپںم€‚ هڈ¤ç”ںن»£م€پ5ه„„4200ن¸‡ه¹´مپ‹م‚‰2ه„„5100ن¸‡ه¹´ه‰چ(Ma) مپ¯م€پمƒ‘مƒ³مƒژمƒ†م‚£م‚¢مپ®ه´©ه£ٹç›´ه¾Œمپ§م€پهœ°çگƒè¦ڈو¨،مپ®و°·و²³وœںمپ®çµ‚م‚ڈم‚ٹمپŒه§‹مپ¾مپ£مپںم€‚هˆوœںمپ®هڈ¤ç”ںن»£م‚’é€ڑمپکمپ¦م€پهœ°çگƒمپ®é™¸ه،ٹمپ¯مپ‹مپھم‚ٹمپ®و•°مپ®و¯”較çڑ„ه°ڈمپ•مپھه¤§é™¸مپ«هˆ†ه‰²مپ•م‚Œمپںم€‚و™‚ن»£مپ®çµ‚م‚ڈم‚ٹمپ«هگ‘مپ‹مپ£مپ¦م€په¤§é™¸مپ¯مƒ‘مƒ³م‚²م‚¢مپ¨ه‘¼مپ°م‚Œم‚‹è¶…ه¤§é™¸مپ«é›†مپ¾م‚ٹم€پهœ°çگƒمپ®é™¸هœ°مپ®ه¤§éƒ¨هˆ†مپŒهگ«مپ¾م‚Œمپ¦مپ„مپں[74]م€‚444Maمپ«ه§‹مپ¾مپ£مپںم‚·مƒ«مƒ«ç´€مپ®é–“[74]م€پم‚´مƒ³مƒ‰مƒ¯مƒٹمپ¯هچ—é«کç·¯ه؛¦مپ«هگ‘مپ‹مپ£مپ¦م‚†مپ£مپڈم‚ٹمپ¨هچ—ن¸‹مپ—ç¶ڑمپ‘مپںم€‚و°·ه† مپ¨و°·و²³مپ®èچ解مپ¯وµ·é¢ن¸ٹوک‡مپ®ن¸€ه› مپ¨مپھمپ£مپںم€‚مپ“مپ®مپ“مپ¨مپ¯م€پم‚·مƒ«مƒ«ç´€مپ®ه †ç©چ物مپŒن¾µé£ںمپ•م‚Œمپںم‚ھمƒ«مƒ‰مƒ“م‚¹ç´€مپ®ه †ç©چ物م‚’覆مپ£مپ¦مپٹم‚ٹم€پن¸چو•´هگˆم‚’ه½¢وˆگمپ—مپ¦مپ„م‚‹مپ¨مپ„مپ†ن؛‹ه®ںمپ‹م‚‰وژ¨ه¯ںمپ§مپچم‚‹م€‚ن»–مپ®م‚¯مƒ©مƒˆمƒ³مپ¨ه¤§é™¸مپ®و–片مپŒèµ¤éپ“ن»کè؟‘مپ§ن¸€ç·’مپ«و¼‚وµپمپ—م€پمƒ¦مƒ¼مƒ©مƒ،مƒھم‚«ه¤§é™¸مپ¨مپ—مپ¦çں¥م‚‰م‚Œم‚‹ç¬¬2مپ®è¶…ه¤§é™¸مپ®ه½¢وˆگم‚’é–‹ه§‹مپ—مپںم€‚  Animation of Pangaea rifting
هœ°çگƒمپ®è،¨é¢مپ¯م€پن½•هچپه„„ه¹´مپ«م‚‚م‚ڈمپںمپ£مپ¦çµ¶مپˆمپڑه½¢م‚’ه¤‰مپˆمپ¦مپچمپ¦مپٹم‚ٹم€په¤§é™¸مپ®ه½¢وˆگم‚„هˆ†è£‚م€پهœ°è،¨م‚’و¨ھهˆ‡م‚‹ç§»ه‹•م€پمپمپ—مپ¦و™‚مپ«مپ¯çµگهگˆمپ—مپ¦è¶…ه¤§é™¸م‚’ه½¢وˆگمپ—مپ¦مپچمپںم€‚çں¥م‚‰م‚Œمپ¦مپ„م‚‹وœ€هڈ¤مپ®è¶…ه¤§é™¸مپ§مپ‚م‚‹مƒمƒ‡م‚£مƒ‹م‚¢مپ¯ç´„10ه„„ه¹´ه‰چمپ«èھ•ç”ںمپ—م€پç´„7ه„„ه¹´ه‰چمپ«هˆ†è£‚مپ—ه§‹م‚پمپںم€‚هˆ†مپ‹م‚Œمپںه¤§é™¸مپ¯ه¾Œمپ«ه†چçµگهگˆمپ—م€پ6ه„„مپ‹م‚‰5ه„„4000 ن¸‡ه¹´ه‰چمپ«مƒ‘مƒ³مƒژمƒ†م‚£م‚¢م‚’ه½¢وˆگمپ—م€پوœ€ه¾Œمپ«2ه„„ه¹´ه‰چمپ«هˆ†è£‚مپ—مپ¦مƒ‘مƒ³م‚²م‚¢مپ«مپھمپ£مپںم€‚ن¸‰ç•³ç´€مپ«مپ¯و·±وµ·ه †ç©چ物مپŒه †ç©چمپ—م€پمپمپ®ه¾Œم€پوµ·و´‹مƒ—مƒ¬مƒ¼مƒˆمپ®و²ˆمپ؟è¾¼مپ؟مپ«م‚ˆمپ£مپ¦و¶ˆه¤±مپ—مپںمپںم‚پم€پن¸‰ç•³ç´€مپ®ه¤–و´‹مپ«مپ¤مپ„مپ¦مپ¯مپ»مپ¨م‚“مپ©çں¥م‚‰م‚Œمپ¦مپ„مپھمپ„م€‚超ه¤§é™¸مƒ‘مƒ³م‚²م‚¢مپ¯ن¸‰ç•³ç´€م€پ特مپ«ن¸‰ç•³ç´€ه¾Œوœںمپ«è£‚مپ‘مپںمپŒم€پمپ¾مپ هˆ†é›¢مپ—مپ¦مپ„مپھمپ‹مپ£مپںم€‚مƒ‘مƒ³م‚²م‚¢مپ®وœ€هˆمپ®هˆ†è£‚م‚’ç¤؛مپ™هœ°و؛ه¸¯مپ®وœ€هˆمپ®éوµ·و´‹ه †ç©چ物مپ¯م€پن¸‰ç•³ç´€ه¾Œوœںمپ®م‚‚مپ®مپ§مپ‚م‚‹[75]م€‚ ن¸‰ç•³ç´€مپ¯è¥؟مƒ¨مƒ¼مƒمƒƒمƒ‘مپ§è‰¯مپڈç ”ç©¶مپ•م‚Œمپ¦مپ„م‚‹م‚‚مپ®مپ®م€پ1 مپ¤مپ®è¶…ه¤§é™¸ه،ٹمپ®وµ·ه²¸ç·ڑمپŒé™گم‚‰م‚Œمپ¦مپ„م‚‹مپںم‚پم€پن¸‰ç•³ç´€مپ®وµ·و´‹ه †ç©چ物مپ¯ن¸–ç•Œçڑ„مپ«و¯”較çڑ„مپ¾م‚Œمپ§مپ‚م‚‹م€‚مپںمپ¨مپˆمپ°م€پهŒ—ç±³مپ§مپ¯م€پوµ·و´‹ه †ç©چ物مپ¯è¥؟هپ´مپ®مپ„مپڈمپ¤مپ‹مپ®éœ²é مپ«é™گه®ڑمپ•م‚Œمپ¦مپ„م‚‹م€‚مپ—مپںمپŒمپ£مپ¦م€پن¸‰ç•³ç´€مپ®ه±¤ه؛ڈمپ¯م€پم‚¨م‚¹مƒ†مƒھم‚¢ç”²و®»é،م‚„陸ç”ںè„ٹو¤ژه‹•ç‰©مپھمپ©مپ®ن¸»مپ«مƒ©م‚°مƒ¼مƒ³م‚„ه،©هˆ†و؟ƒه؛¦مپ®é«کمپ„ç’°ه¢ƒمپ«ç”ںوپ¯مپ™م‚‹ç”ں物مپ«هں؛مپ¥مپ„مپ¦مپ„م‚‹[76]م€‚  Geologic time represented by a geological clock, showing the relative lengths of the eons of Earth's history and noting major events  ç”ں物و“¾ن¹±مپ®مƒ‘م‚؟مƒ¼مƒ³م‚„ç—•è·،مپ¯م€پçں³هŒ–مپ—مپںه²©çں³مپ«ن؟هکمپ•م‚Œمپ¦مپ„م‚‹م€‚مپ“مپ®م‚ˆمپ†مپھمƒ‘م‚؟مƒ¼مƒ³مپ®ç ”究مپ¯ichnology م€پمپ¾مپںمپ¯م€Œç—•è·،هŒ–çں³م€چمپ®ç ”究مپ¨ه‘¼مپ°م‚Œم‚‹م€‚مپ“م‚Œمپ¯م€پç”ں物و“¾ن¹±ç¨®مپ®ه ´هگˆم€په‹•ç‰©مپŒç©´م‚’وژکمپ£مپںم‚ٹهں‹مپ¾مپ£مپںم‚ٹمپ™م‚‹مپ“مپ¨مپ«م‚ˆمپ£مپ¦و®‹مپ•م‚ŒمپںهŒ–çں³مپ§مپ‚م‚‹م€‚مپ“م‚Œمپ¯م€پمپ“م‚Œم‚‰مپ®ه‹•ç‰©مپŒو®‹مپ—مپں足è·،مپ¨و¯”較مپ™م‚‹مپ“مپ¨مپŒمپ§مپچم‚‹م€‚ه ´هگˆمپ«م‚ˆمپ£مپ¦مپ¯م€پç”ں物و“¾ن¹±مپŒéه¸¸مپ«و™®هڈٹمپ—مپ¦مپ„م‚‹مپںم‚پم€پç©چه±¤ه±¤م‚„م‚¯مƒم‚¹مƒ™مƒƒمƒ‰مپھمپ©مپ®ه †ç©چو§‹é€ مپŒه®Œه…¨مپ«و¶ˆو»…مپ™م‚‹م€‚مپ—مپںمپŒمپ£مپ¦م€پمپم‚Œمپ¯هœ°è³ھه¦ه†…مپ®ه †ç©چه¦مپ¨ه±¤ه؛ڈه¦مپ®هˆ†é‡ژمپ«ه½±éں؟م‚’ن¸ژمپˆم‚‹م€‚ç”ں物و“¾ن¹±ç¨®مپ®ichnofabrics مپ®ç ”究مپ§مپ¯م€پهŒ–çں³مپ®و·±مپ•م€پهŒ–çں³مپ®و¨ھو–é¢م€پمپٹم‚ˆمپ³هŒ–çں³مپ®وکژç¢؛مپ• (مپ™مپھم‚ڈمپ،م€پمپ©مپ®ç¨‹ه؛¦وکژç¢؛مپ«ه®ڑ義مپ•م‚Œمپ¦مپ„م‚‹مپ‹)مپھمپ©مپ®وƒ…ه ±م‚’هˆ©ç”¨مپ—مپ¦م€پهڈ¤مپ„ه †ç©چ物مپ§ç™؛ç”ںمپ—مپںو´»ه‹•م‚’è©•ن¾،مپ™م‚‹[77]م€‚é€ڑه¸¸م€پهŒ–çں³مپŒو·±مپ‘م‚Œمپ°و·±مپ„مپ»مپ©م€پو¨™وœ¬مپ¯م‚ˆم‚ٹم‚ˆمپڈن؟هکمپ•م‚Œم€پوکژç¢؛مپ«ه®ڑ義مپ§مپچم‚‹[77]م€‚ و½®و±گم€پو²؟ه²¸مپٹم‚ˆمپ³و·±وµ·مپ®ه †ç©چ物مپ‹م‚‰مپ®وµ·و´‹ه †ç©چ物مپ«مپ¯م€پç”ں物و“¾ن¹±مپ®é‡چè¦پمپھه¾®é‡ڈهŒ–çں³مپŒç™؛見مپ•م‚Œمپ¦مپ„م‚‹م€‚مپ•م‚‰مپ«م€پç ‚ن¸کم€پمپ¾مپںمپ¯é¢¨وˆگمپ®ه †ç©چ物مپ¯م€په¤ڑ種ه¤ڑو§کمپھهŒ–çں³م‚’ن؟هکمپ™م‚‹مپںم‚پمپ«é‡چè¦پمپ§مپ‚م‚‹[78]م€‚ç”ں物و“¾ن¹±مپ®è¨¼و‹ مپ¯م€پé•·مپ„è¨ک録م‚’هگ«م‚€و·±وµ·ه †ç©چ物م‚³م‚¢مپ§ç™؛見مپ•م‚Œمپ¦مپ„م‚‹مپŒم€پم‚³م‚¢م‚’وٹ½ه‡؛مپ™م‚‹è،Œç‚؛مپ¯م€پ特مپ«وµ…مپ„و·±مپ•مپ§ç”ں物ن¹±وµپمپ®ه…†ه€™م‚’ن¹±مپ™هڈ¯èƒ½و€§مپŒمپ‚م‚‹[79]م€‚特مپ«ç¯€è¶³ه‹•ç‰©مپ¯م€پ風وˆگه †ç©چ物مپ®ç”ں物ن¹±وµپمپ®هœ°è³ھه¦çڑ„è¨ک録مپ«مپ¨مپ£مپ¦é‡چè¦پمپ§مپ‚م‚‹م€‚ç ‚ن¸کمپ®è¨ک録مپ¯م€پن¸ç”ںن»£مپ®ن¸‹éƒ¨م€پ250 Ma مپ¾مپ§مپ•مپ‹مپ®مپ¼م‚‹ه‹•ç‰©مپ®ç©´وژکم‚ٹمپ®ç—•è·،م‚’ç¤؛مپ—مپ¦مپ„م‚‹[78]مپŒم€پن»–مپ®ه †ç©چ物مپ«مپٹمپ‘م‚‹ç”ں物و“¾ن¹±مپ¯ 550Ma مپ¾مپ§مپ•مپ‹مپ®مپ¼مپ£مپ¦è¦‹م‚‰م‚Œمپ¦مپ„م‚‹[80][81]م€‚ ç ”ç©¶هڈ²و·±وµ·ه †ç©چ物مپ«é–¢مپ™م‚‹وœ€هˆمپ®ن¸»è¦پمپھç ”ç©¶مپ¯م€پ1872ه¹´مپ‹م‚‰ 1876ه¹´مپ«مپ‹مپ‘مپ¦HMSمƒپمƒ£مƒ¬مƒ³م‚¸مƒ£مƒ¼éپ ه¾پéڑٹمپ«م‚ˆمپ£مپ¦è،Œم‚ڈم‚Œم€پوµ·و°´مپ¨وµ·و´‹ه †ç©چ物م‚’م‚µمƒ³مƒ—مƒھمƒ³م‚°مپ™م‚‹مپںم‚پمپ«ç´„ 70,000 وµ·é‡Œم‚’移ه‹•مپ—مپں[82]م€‚éپ ه¾پمپ®ç§‘ه¦çڑ„ç›®و¨™مپ¯م€پمپ•مپ¾مپ–مپ¾مپھو·±مپ•مپ§وµ·و°´مپ®ç‰©çگ†çڑ„و¸¬ه®ڑم‚’è،Œمپ†مپ“مپ¨مپ¨م€پهکهœ¨مپ™م‚‹ç²’هگçٹ¶ç‰©è³ھم‚„وµ·و´‹ç”ں物مپ¨مپ¨م‚‚مپ«هŒ–ه¦çµ„وˆگم‚’و±؛ه®ڑمپ§مپچم‚‹م‚ˆمپ†مپ«م‚µمƒ³مƒ—مƒ«م‚’وژ،هڈ–مپ™م‚‹مپ“مپ¨مپ§مپ‚مپ£مپںم€‚مپ“م‚Œمپ«مپ¯م€پو·±وµ·ه؛•مپ‹م‚‰مپ®م‚µمƒ³مƒ—مƒ«مپ®وژ،هڈ–مپ¨ه †ç©چ物مپ®هˆ†وگمپŒهگ«مپ¾م‚Œمپ¦مپ„مپں[83]م€‚مƒپمƒ£مƒ¬مƒ³م‚¸مƒ£مƒ¼هڈ·مپ®èˆھوµ·ن»¥ه‰چمپ¯م€پوµ·و´‹ه¦مپ¯ن¸»مپ«وٹ•و©ںçڑ„مپ§مپ‚مپ£مپں[82]م€‚هˆم‚پمپ¦مپ®çœںمپ®وµ·و´‹م‚¯مƒ«مƒ¼م‚؛مپ¨مپ—مپ¦م€پمƒپمƒ£مƒ¬مƒ³م‚¸مƒ£مƒ¼éپ ه¾پمپ¯ه¦è،“مپٹم‚ˆمپ³ç ”究هˆ†é‡ژه…¨ن½“مپ®هں؛ç¤ژم‚’築مپ„مپں[84]م€‚ ه¤§é™¸ç§»ه‹•مپ®هˆوœںمپ®çگ†è«–مپ§مپ¯م€پéپ‹ه‹•ن¸مپ®ه¤§é™¸مپŒه›؛ه®ڑمپ•م‚Œمپںن¸چه‹•مپ®وµ·ه؛•م‚’م€Œوژ»مپچهˆ†مپ‘مپںم€چ(plowed)مپ¨è،¨çڈ¾مپ•م‚Œمپںم€‚ 1960ه¹´ن»£ه¾Œهچٹمپ«م€پوµ·ه؛•è‡ھن½“مپŒç§»ه‹•مپ—م€پن¸ه¤®مپ®مƒھمƒ•مƒˆè»¸مپ‹م‚‰ه؛ƒمپŒم‚‹مپ¨مپچمپ«ه¤§é™¸م‚’éپ‹مپ¶مپ¨مپ„مپ†è€ƒمپˆمپŒم€پ Harold Hessمپ¨Robert Dietzمپ«م‚ˆمپ£مپ¦وڈگو،ˆمپ•م‚Œمپں[85][86]م€‚مپ“مپ®çڈ¾è±،مپ¯ن»ٹو—¥م€پمƒ—مƒ¬مƒ¼مƒˆمƒ†م‚¯مƒˆمƒ‹م‚¯م‚¹مپ¨مپ—مپ¦çں¥م‚‰م‚Œمپ¦مپ„م‚‹م€‚ 2مپ¤مپ®مƒ—مƒ¬مƒ¼مƒˆمپŒé›¢م‚Œمپ¦مپ„م‚‹ه ´و‰€م€پن¸ه¤®وµ·ه¶؛مپ§مپ¯م€پوµ·ه؛•مپŒه؛ƒمپŒم‚‹éپژ程مپ§و–°مپ—مپ„وµ·ه؛•مپŒçµ¶مپˆمپڑه½¢وˆگمپ•م‚Œم‚‹[87]م€‚1968ه¹´م€پوµ·و´‹èھ؟وں»èˆ¹Glomar ChallengerمپŒé€²و°´مپ—م€پ15ه¹´é–“مپ«م‚ڈمپںم‚‹و·±وµ·وژکه‰ٹ計画مپ«ç€و‰‹مپ—مپںم€‚مپ“مپ®مƒ—مƒم‚°مƒ©مƒ مپ¯م€پن¸ه¤®وµ·ه¶؛مپ‹م‚‰éپ مپ„مپ»مپ©ه²©çں³مپŒهڈ¤مپ„مپ“مپ¨م‚’ç¢؛èھچمپ™م‚‹ه²©çں³م‚µمƒ³مƒ—مƒ«م‚’هڈژ集مپ™م‚‹مپ“مپ¨مپ«م‚ˆم‚ٹم€پوµ·ه؛•و‹،ه¤§ن»®èھ¬م‚’و”¯وŒپمپ™م‚‹é‡چè¦پمپھمƒ‡مƒ¼م‚؟م‚’وڈگن¾›مپ—مپں[88][89]م€‚ هڈ‚考و–‡çŒ®
م‚½مƒ¼م‚¹
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




.jpg)
,_Haeckel_(28187768550).jpg)









.jpg)


![Sediment supply from terrigenous and biological sources as well as its dispersion and settling through the water columnâ€ٹ[28]](%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Ocean_hemipelagic_and_pelagic_processes.webp)

_current.webp)









_Various_diatoms.jpg)




.jpg)
.jpg)


Portal di Ensiklopedia Dunia













