Сауроподи
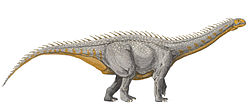
Сауроподи, чии членови се познати како сауроподи[2] (од sauro- + -pod“, гуштер“) — клад од сауриски диносауруси. Сауроподите имале многу долг врат, долги опашки, мали глави (во однос на остатокот од нивното тело) и четири дебели нозе како столбови. Тие се познати по огромните големини што ги достигнале некои видови, а групата ги вклучува најголемите животни што некогаш живееле на копно. Добро познати родови вклучуваат Брахиосаурус, Дилподокус, Апатосаурус и Бронтосаурус.[3][4] Најстарите познати недвосмислени диносауруси од сауропод се познати уште од раната јура.[5] Исаносаурус и Антетонитрус првично биле опишани како сауроподи од периодот на тријас,[6][7] но нивната возраст, а во случајот и на Антетонитрус, исто така, и неговиот статус на сауропод, потоа биле доведени во прашање.[8][9] Сауроподоморфните патеки слични на Сауропода од формацијата Флеминг Фјорд (Гренланд) може, сепак, да укажуваат на појава на групата во доцниот тријас. Од крајот на јурата (пред 150 милиони години), сауроподите станале широко распространети (особено диплодоцидите и брахиосауридите). Во Доцната Креда, една група сауроподи, титаносаури, ги заменила сите други и имале скоро глобална распространетост. Како и да е, како и со сите други не-птичји диносауруси, живи во тоа време, титаносаурусите изумреле во настанот на исчезнување креда-палеоген. Фосилизирани остатоци од сауроподи биле пронајдени на секој континент, вклучително и на Антарктикот.[10][11][12][13] Називот „Сауропода“ е смислен од Отниел Марш во 1878 година и потекнува од грчки, што значи „стапало на гуштер“.[14] Сауроподите се една од најпрепознатливите групи на диносауруси и станале термин во популарната култура поради нивната импресивна големина. Комплетни наоди од фосили на сауроподи се ретки. Многу видови, особено најголемите, се познати само од изолирани и одделени коски. На многу скоро комплетни примероци им недостасуваат глави, врвови на опашката и екстремитети. ОписСауроподите биле тревојади (растителни), обично со прилично долг врат [15] четириножни (четири нозе), често со запчести (во форма на шпатула: широки на врвот, тесни на вратот) заби. Тие имале мали глави, масивни тела, а повеќето имале долги опашки. Нивните задни нозе биле дебели, прави и моќни, завршувајќи на стапалата со пет прсти, иако само внатрешните три (или во некои случаи и четири) имале канџи. Нивните предни екстремитети биле прилично витки и обично завршувале со раце слични на столб, изградени за поддршка на тежината; честопати само палецот имал канџи. На многу илустрации на сауроподи во телото им недостасуваат овие факти, неточно прикажани сауроподи со копита кои ги затвораат нозете без канџи или повеќе од три канџи или копита на рацете. Проксималните каудални пршлени се крајно дијагностички откриени за сауроподите.[16] Големина Најдефинирачката одлика на сауроподите била нивната големина. Дури и џуџестите сауроподи (можеби од 5 до 6) метри, или 20 долги стапки) биле сметани меѓу најголемите животни во нивниот екосистем . Нивните единствени вистински конкуренти според големината биле китовите, како синиот кит . Но, за разлика од китовите, сауроподите биле првенствено копнени животни . Нивната структура на телото не се разликувала многу како другите диносауруси, можеби поради ограничувањата во големината, но тие покажале голема разновидност. Некои, како диплоодоцидите, имале неверојатно долги опашки, кои можеби биле во можност да ги користат како камшик како сигнал или да ги одвратат или повредат грабливците,[17] или да направат звучен ѕид .[18][19] Суперсаурус, на 33-34 метри долг,[20] бил најдолгиот сауропод познат од разумно комплетни остатоци, но и други, како стариот рекордер, Диплодокус, биле исто така долги. Холотипот (и сега изгубен) пршлен на Amphicoelias fragillimus (сега Марапунисаурус) можеби потекнувало од животно долго 58 метри;[21] неговата вертебрална колона би била значително подолга од онаа на синиот кит. Сепак, истражувањето објавено во 2015 година претпоставува дека проценките за големината на A. fragillimus можеби биле претерани.[22] Најдолгиот познат диносаурус од разумен фосилен материјал е веројатно Аргентиносаурусот хуинкуленсис со проценка на должината од 35 до 36 метри според најновите истражувања.[23][24] Сепак, гигантскиот примерок од Баросаурус BYU 9024 можеби бил уште поголем, достигнувајќи должина од 45-48 метри (148-157 стапки).[25] Најдолгото копнено животно што живее денес, мрежестиот питон достигнува должина од само 6,95 метри [26] Другите, како брахиосауридите, биле екстремно високи, со високи рамена и екстремно долги вратови. Највисокиот сауропод бил примерок од гигантот Баросаурус со височина од 22 метри.[23] За споредба, жирафата, највисоката од сите живи копнени животни, има само 4,8 - 5,5 метри височина. Најдобар доказ укажува на тоа дека најмасивни биле Аргентиносаурусите (65-80 метрички тони [23][24][27] ), Маменчисаурусите (60-80 метрички тони), гигантскиот примерок од Баросаурус (60-80 + метрички тони [25] ) и Патаготитанот со Пуертасаурусот (50-55 метрички тони). Имало и слаби (и сега исчезнати) докази дека таканаречениот Брухаткајосаурус можеби тежел над 175 метрички тони, но ова е доведено во прашање.[28] Наводната фибула е веројатно фемур на животно нешто поголемо од Драдноутус . Тежината на Amphicoelias fragillimus се проценува на 122,4 метрички тони [21] но истражувањето од 2015 година тврди дека овие проценки можеби биле многу претерани.[22] Дополнителни откритија укажуваат на голем број видови кои веројатно достигнале или надминале тежина од 40 тони.[29] Најголемото копнено животно живо денес, грмушкиот слон, тежи не повеќе од 10,4 метрички тони .[30] Меѓу најмалите сауроподи биле примитивниот Омденосаурус (4 метри висок со 13 метри должина), џуџестиот титаносаурус Магјаросаурус (6 метри должина), , и џуџестиот брахиосаурид Европасаурус, (6,2 метри должина).[31] Неговиот мал раст веројатно бил резултат на изоларно џуџест потег кај популација на сауроподи изолирани на остров на доцната јура во денешната област Лангенберг во северна Германија.[32][33] Диплодоцидот сауропод Брахитрахелопан бил најкраткиот член во својата група поради невообичаено краткиот врат. За разлика од другите сауроподи, чиј врат можел да порасне и до четири пати поголем од грбот, вратот на Брахитрахелопан бил пократок од ’рбетот. Фосили од можеби најголемиот диносаурус некогаш пронајдени се откриени во 2012 година во провинцијата Неукен, северозападна Патагонија, Аргентина. Се верува дека тие потекнуваат од титаносаурус, кои биле меѓу најголемите сауроподи.[34][35] На или малку пред 29 март 2017 година, трага од сауропод долг околу 1,6 метри (1,7 метри) била пронајдена во Валмадани, во регионот Кимберли во Западна Австралија.[36] Во извештајот се вели дека тоа е досега најголемото познато. Во 2020 година, Молина-Перес и Лараменди ја процениле големината на животното на 31 метри (102 стапки) и 72 тони (79,4 кратки тони) врз основа на долгото стапало од 1,75 метри (5,7 стапки).[23] Екстремитети и стапалаКако масивни четириножни животни, сауроподите развиле специјализирани екстремитети на гравипорталната (товарна) тежина. Задните нозе биле широки и задржувале три канџи кај повеќето видови.[37] Особено невообичаени во споредба со другите животни биле високо модифицираните предни стапала (манус). Предните стапала на сауроподите биле многу различни од оние на современите големи четириножци, како што се слоновите. Коските на раката биле наредени во целосно вертикални колони, со крајно редуцирани коскени прсти (и покрај тоа што не е јасно дали најпримитивните сауроподи, како што Вулканодон и Барапасаурус, имале таква претходница).[38] Предните стапала биле толку модифицирани во евздуроподи, што поединечни цифри не би биле видливи во животот. Распоредот на колоните на коските на предните нозе (метакарпалните ) бил полукружен, така што отпечатоците од предните нозе на сауропод се во форма на потковица. За разлика од слоновите, отпечатените докази покажуваат дека на сауроподите немало никакво обложување за да ги поддржат предните стапала, што ги прави вдлабнати.[38] Единствената канџа видлива кај повеќето сауроподи била карактеристичниот нокт на палецот (поврзан со цифрата I). Речиси сите сауроподи имале таква канџа, иако не е позната со каква цел служела. Ноктот бил најголем (како и висок и странично зарамнет) кај диплодоцидите и многу мал кај брахиосауридите, од кои некои се чини дека целосно го изгубиле ноктот врз основа на доказите.[39] Титаносаурусите можеби го изгубиле ноктот на палецот целосно (со исклучок на раните форми, како што е Јаненшија ). Титаносаурусите биле најневообичаени меѓу сауроподите, бидејќи покрај надворешната канџи, тие целосно ги изгубиле прстите на предната нога. Напредните титаносауруси немале прсти, и оделе само на „трупците“ во форма на потковица, составени од колонарните метакарпални коски.[40] Отпечатените докази од Португалија покажуваат дека, барем кај некои сауроподи (веројатно брахиосауриди), дното и страните на колоната на предните нозе најверојатно биле покриени со мали, боцкави скали, што оставале отпечатоци.[41] Кај титаносаурусите, краевите на метакарпалните коски кои допирале со земјата биле невообичаено широки и квадратни, а некои примероци ги зачувуваат остатоците од мекото ткиво што ја покриваат оваа област, што укажува на тоа дека предните нозе биле обложени.[40] Метју Бонан [42][43] покажал дека долгите коски на диносаурусот од сауропод растеле изометрично, односно немало многу или воопшто не се менувала во формата, бидејќи малолетни сауроподи станале гигантски возрасни. Бонан сугерирал дека овој необичен модел на скалирање (повеќето ’рбетници покажуваат значителни промени во обликот на долгите коски поврзани со зголемување на поддршката на телесната тежина) може да биде поврзан со принципот на stilt-walker (предложен од аматерскиот научник Џим Шмит) во кој им дозволувале долгите нозе на возрасните сауроподи за лесно покривање на големи растојанија без промена на нивната целокупна механика. Воздушни вреќиЗаедно со другите сауриски диносауруси (како што се тероподи, вклучително и птици), сауроподите имале систем на воздушни вреќи, докажано со вдлабнатините и шуплините во повеќето од нивните пршлени. Пневматски, шупливи коски се карактеристична црта на сите сауроподи.[44] Овие воздушни простори ја намалувале вкупната тежина на масивните вратови што ги имале сауроподите, и системот на воздушни вреќи, воопшто, овозможувајќи проток на воздух во еден правец низ беспомошни бели дробови, им овозможувало на сауроподите да добијат доволно кислород.[45] Дупчењето на коските на сауропод како птица било препознаено на почетокот на студијата за овие животни и, всушност, барем еден примерок од сауропод пронајден во 19 век (Орнитопсис) првично бил погрешно идентификуван како птеросаурус (летачки влекач).[46] Оклоп Некои сауроподи имале оклоп. Имале родови со мали шипки на опашот, како Шуносаурус, и неколку титаносаури, како што се Салтасаурус и Ампелосаурус, имале мали коскени остеодерми кои покривале делови од нивните тела. ЗабиСтудија на Мајкл Д'Емик и неговите колеги од Универзитетот Стони Брук открила дека сауроподите еволуирале високи стапки на заби за да бидат во чекор со нивните големи апетити. Студијата сугерира дека Нигерсаурус, на пример, го заменувал секој заб на секои 14 дена, Камарасаурус го заменувал секој заб на секои 62 дена, а Диплодокус го заменувал секој заб еднаш на секои 35 дена.[47] Научниците откриле дека квалитетите на забот влијаат на тоа колку време требало за да расте новиот заб. На забите на Камарасаурус им требало подолго време да пораснат од оние за Дилодокусот бидејќи биле поголеми.[48] Исто така, било забележано од Д'Емик и неговиот тим дека разликите помеѓу забите на сауроподите, исто така, укажуваат на разлика во исхраната. Диплодокусот јадел растенија ниско до земјата а Камарасаурусот прелистувал лисја од горните и средните гранки. Според научниците, специјализирањето на нивните диети им помогнало на различните тревојади диносауруси да коегзистираат.[47][48] ВратВратовите на сауроподите биле над 15 метри долги, цели шест пати подолги од светскиот рекорд на вратот на жирафата.[45] Овозможувањето на ова било голем број на основни физиолошки одлики. Целокупната голема димензија на телото на диносаурусите и четиридадечкиот став обезбедувале стабилна основа за поддршка на вратот, а главата била еволуирана да биде многу мала и лесна, губејќи ја можноста за орална обработка на храната. Смалувајќи ги главите на едноставни алатки за берба кои ги внесувале растенијата во телото, на сауроподите им требало помалку енергија за да ги кренат главите и на тој начин биле во можност да развијат вратови со помалку густи мускули и сврзно ткиво. Ова драстично ја намалило вкупната маса на вратот, овозможувајќи понатамошно издолжување. Сауроподите исто така имале голем број на адаптации во нивната скелетна структура. Некои сауроподи имале дури 19 вратни пршлени, додека скоро сите цицачи се ограничени на само седум. Покрај тоа, секој пршлен бил исклучително долг и имал голем број празни места во нив кои биле исполнети само со воздух. Систем со воздушни вреќички поврзан со просторите не само што ги осветлувало долгите вратови, туку ефикасно го зголемувал протокот на воздух низ душникот, помагајќи им на суштествата да дишат доволно воздух. Со развивање на пршлени кои се состојат од 60% воздух, сауроподите биле во можност да ја минимизираат количината на густа, тешка коска без да ја жртвуваат можноста да земат доволно големи вдишувања за да го напојат целото тело со кислород.[45] Според Кент Стивенс, компјутерски моделирани реконструкции на скелети направени од пршлени укажуваат на тоа дека вратовите на сауроподите биле способни да избришат големи области за хранење без потреба да ги движат нивните тела, но не биле во можност да бидат повлечени во позиција многу над рамената за да ја истражат областа.[49] Друга предложена функција на долгите вратови на сауроподите во суштина бил радијатор за справување со екстремната количина на топлина произведена од нивната голема телесна маса. Имајќи предвид дека метаболизмот вршел огромна работа, сигурно генерирал и голема количина на топлина, а елиминирањето на овој вишок топлина би бил од суштинско значење за опстанокот.[50] Исто така, било предложено дека долгите вратови би ги ладеле вените и артериите што одат кон мозокот, избегнувајќи претерано загреана крв да стигне до главата. Всушност било откриено дека зголемувањето на метаболичката стапка како резултат на вратот на сауроподите била нешто повеќе од компензирано со дополнителната површина од која топлината може да се распрсне.[51] ПалеобиологијаЕкологијаКога за прв пат биле откриени сауроподи, нивната огромна големина натерала многу научници да ги споредат со денешните китови. Повеќето студии во 19 и почетокот на XX век заклучиле дека сауроподите биле преголеми за да ја издржат нивната тежина на копно и затоа морале да биле главно водни. Повеќето животни реставрации на сауроподи во уметноста низ првите три четвртини од 20 век ги прикажале целосно или делумно потопени во вода.[52] Овој ран поим бил ставен под сомнеж, започнувајќи во 50-тите години на минатиот век, кога студија на Кермак (1951) покажала дека, ако животното се потопи во неколку метри вода, притисокот би бил доволен за фатално пропаѓање на белите дробови и дишните патишта.[53] Сепак, ова и другите рани студии за екологијата на сауропод имале недостатоци во тоа што тие игнорирале значителен број докази дека телата на сауроподите биле претежно проникнати во воздушни вреќи . Во 1878 година, палеонтологот Коуп дури ги нарекол овие структури како да „пловат“. Почнувајќи од 1970-тите, започнале да се истражуваат ефектите на воздушните вреќи на сауроподите врз нивниот наводен воден начин на живот. Палеонтолозите како Кумбс и Бакер го користеле ова, како и докази од седиментологијата и биомеханиката, за да покажат дека сауроподите биле првенствено копнени животни. Во 2004 година, Д.М. Хендерсон забележале дека, поради нивниот широк систем на воздушни вреќи, сауроподите би биле подвижни и немало да можат да ги потопат нивните торза целосно под површината на водата; со други зборови, тие би лебделе и немало да бидат во опасност од колапс на белите дробови поради притисокот на водата при пливање.[52] Доказите за пливање во сауроподи доаѓаат од фосилни патеки за кои повремено се открива дека ги зачувуваат само впечатоците од предниот дел (манусот). Хендерсон покажал дека таквите патеки може да се објаснат со сауроподи со долги предни екстремитети (како што се макронарците ) кои лебдат во релативно плитка вода доволно длабоко за да ги задржат пократките задни нозе од долниот дел и да ги користат предните екстремитети за да ловат напред.[52] Сепак, поради нивните пропорции на тело, пловечките сауроподи исто така би биле многу нестабилни и лошо прилагодени за подолги периоди во водата. Овој начин на водено движење, во комбинација со неговата нестабилност, го натерало Хендерсон да ги опише сауроподите во вода како „вртоглави ловџии“. Иако сауроподите не можеле да бидат водни како што биле историски прикажани, постојат докази дека тие претпочитаат влажни и крајбрежни живеалишта. Стапалата на сауроподите обично се наоѓаат по крајбрежјето или преминуваат поплавни рамнини, а фосилите на сауропод често се наоѓаат во влажни средини или се мешаат со фосили на морски организми.[52] Добар пример за ова би биле остатоците во наоѓалиштата на лагуната на шкотскиот остров Скај .[54] Родителска грижа Многу редови докази на фосили,, укажуваат на тоа дека сауроподите биле животни кои формирале стада . Сепак, составот на стадата се разликувал помеѓу видовите. Некои коскени корита (Bone bed), на пример, место од средната јура во Аргентина, се чини дека покажуваат стада составени од лица од различни возрасни групи, мешајќи малолетници и возрасни. Сепак, голем број други фосилни места и патеки покажуваат дека многу видови на сауроподи патувале во стада одделени според возраста, а малолетниците формирале стада одделени од возрасните. Таквите одделени стратегии за пасење се пронајдени кај видови како што се Аламосаурус, Белусаурус и некои диплодоциди.[55] Во прегледот на доказите за различни типови стада, Мајерс и Фиорило се обиделе да објаснат зошто се чини дека сауроподите често формирале одделени стада. Студиите за микроскопско трошење на забите покажуваат дека малолетни сауроподи имале исхрана која се разликувала од нивните возрасни колеги, па затоа пасењето заедно немало да биде толку продуктивно како што се одгледувало одделно, каде што одделни членови на стадото можеле да се хранат на координиран начин. Огромната разлика во големината помеѓу малолетниците и возрасните исто така може да учествувала во различните стратегии за хранење.[55]  Бидејќи сегрегацијата на малолетници и возрасни мора да се случила наскоро по изведувањето, Мајерс и Фиорило заклучиле дека возрасните можеби не покажувале многу голема родителска грижа.[55] Од друга страна, научниците кои проучувале стада сауроподи мешани по возраст, сугерирале дека овие видови можеби се грижеле за своите млади подолг временски период пред младите да достигнат зрелост.[56] Една студија од 2014 година сугерира дека времето од положување на јајцето до времето на изведување веројатно било од 65 до 82 дена.[57] Не е познато точно колку се разликува сегрегираното наспроти возрасното пасење во различни групи сауроподи. Можеби треба да се откријат дополнителни примери на григорозно однесување од повеќе видови сауроподи за да се откријат повеќе податоци. Став Уште од почетокот на историјата на нивното истражување, научниците, како Озборн, шпекулираат дека сауроподите можеле да се држат на задните нозе, користејќи ја опашката како трета „нога“.[58] Скелетни монтажи со ликот на диплодоцидот Barosaurus lentus го прикажуваат диносаурусот подигнат на задните нозе, кој модел се наоѓа во Американскиот музеј за природна историја. Во труд од 2005 година, Ротшилд и Молнар образложиле дека доколку сауроподите на моменти прифатиле бипедално држење, би имало докази во „рацете“ на предниот екстремитет. Сепак, ниту еден не бил пронајден откако прегледале голем број скелети на сауропод.[59]  Хајнрих Малисон (во 2009 година) бил првиот кој го проучувал физичкиот потенцијал за разни сауроподи да се вратат во триподен став. Малисон открил дека некои ликови претходно поврзани со адаптации за одгледување всушност не биле поврзани (како што се широко поставените коски на колкот на титаносаурусите ) или би го попречувале одгледувањето. На пример, титаносаурусите имале невообичаено флексибилен ’рбет, што би ја намалило стабилноста на триподелното држење и би ги оптоварело повеќе мускулите. Исто така, малку е веројатно дека брахиосауридите би можеле да се држат на задните нозе, бидејќи нивниот центар на гравитација бил далеку понапред од другите сауроподи, што би предизвикало таквиот став да биде нестабилен.[60] Диплодоцидите, од друга страна, се чини дека биле добро прилагодени за триподен став. Диплодоцидите имале центар на маса директно над колковите, давајќи им поголема рамнотежа на двете нозе. Диплодоцидите исто така имале и најподвижни вратови на сауроподи, добро мускулест карличен појас и пршлени на опашката со специјализирана форма што ќе и овозможи на опашката да носи тежина на точката што ја допирала со земјата. Малисон заклучил дека диплодоцидите биле подобро прилагодени на одгледувањето отколку слоновите, кои тоа го прават повремено во дивината,[59][60] Држење на главата и вратот Постои полемика околу тоа како сауроподите ги држеле главата и вратот и положбите што би можеле да ги постигнат во животот. Различни истражувања кои го разгледуваат проблемот од аспекти, како неутрална артикулација на пршлен на вратот и проценка на опсегот на движење, метаболички и енергетски потреби од неверојатно долги вратови и споредба со живи животни, дошле до различни заклучоци. Тврдењето дека долгите вратови на сауроподи биле користени за прелистување на високи дрвја е доведено во прашање врз основа на пресметките за потребната енергија за создавање на артериски крвен притисок за главата доколку се држи исправено.[61] Овие пресметки сугерираат дека ова би зафаќало приближно половина од неговиот внес на енергија.[62] Понатаму, за снабдување со крв во главата, вертикално крената, би бил потребен крвен притисок од околу 700 mmHg (= 0,921 бари ) во срцето. За ова би биле потребни срца 15 пати поголеми од срцата на китовите со слична големина.[63] Ова се користи за да се тврди дека поверојатно е дека долгиот врат обично се држел хоризонтално за да им се овозможи да се хранат со растенија на многу широко подрачје без потреба да ги преместуваат своите тела - потенцијално голема заштеда на енергија за 30 до 40 тони животни. Како поддршка на ова, реконструкциите на вратот на Диплодокус и Апатосаурус покажуваат дека тие се во основа исправени со нежен пад, ориентирајќи ги главата и вратот во „неутрално, неизмастено држење на телото“.[64] Сепак, истражувањето на живи животни тврди дека повеќето живи тетраподи вообичаено ја креваат основата на вратот кога се будни. Инференција од коските за „неутрални положби“, што укажува на поголема хоризонтална положба,[64] може да биде несигурна.[65][66] Истражувањето објавено во 2013 година, кое ги проучувало вратовите на нојот, сепак ја довело во сомнеж проценетата флексибилност на вратот на сауроподот. Студиите на Метју Кобли и сор. откриваат, користејќи компјутерско моделирање, дека мускулните прицврстувања и 'рскавицата присутни во вратот веројатно ќе ја ограничат флексибилноста до значителен степен. Авторите предупредувале да не се проценува опсегот на движење од само користење коски. Ова откритие исто така открива дека сауроподите можеби требало да ги преместат целите тела за да имаат подобар пристап до областите каде што можат да пасат и да ја разгледуваат растителноста.[67][68][69] Патеки и движење   Патеките на движење на сауроподите и другите фосилни стапалки (познати како „ихнити“) се познати од изобилството докази присутни на повеќето континенти. Ихнитите помогнале во поддршката на другите биолошки хипотези за сауроподите, вклучително и општата анатомија на предниот и задниот дел од стапалото (види екстремитети и стапала погоре). Општо земено, отпечатоците од предната нога се многу помали од задните стапала и често се во форма на полумесечина. Повремено ихнитите зачувуваат траги од канџи и помагаат да се потврди кои групи на сауроподи изгубиле канџи или дури делови на предната нога.[70] Патеките на движење на сауроподите од формацијата Вилар дел Арзобиспо во Шпанија го поддржуваат григорозното однесување на групата. Патеките се можеби послични на Sauropodichnus giganteus од која било друга ихногенера, иако се претпоставува дека се од базална титаносауриформа. Патеките се широко распространети, а групацијата близу до Sauropodichnusморфологијата на манусот е во облик на грав и морфологијата на песот е суптриаголна. Не може да се идентификува дали стапалата на стадото биле предизвикани од малолетници или возрасни, поради недостаток на претходна поединечна идентификација на возраста.[71]  Општо, патеките се поделени во три категории врз основа на растојанието помеѓу спротивните екстремитети: тесен мерач, среден мерач и широк мерач. Мерачот на патеката може да помогне да се утврди колку биле широко поставени екстремитетите на разни сауроподи и како тоа може да влијаело на начинот на нивното одење.[70] Студија од Деј и колегите од 2004 година открила дека може да се најде општа шема меѓу групи напредни сауроподи, при што секое семејство на сауроподи се одликува со одредени мерачи на патеки. Откриле дека повеќето сауроподи, освен титаносаурусите, имаат екстремитети со тесен мерач, со силни впечатоци на големите канџи на палецот на предната нога. Патеки со среден мерач со впечатоци на ноктите на предниот дел веројатно припаѓаат на брахиосауридите и други примитивни титаносаурируми, кои се развивале со пошироко поставени екстремитети, но ги задржувале своите канџи. Примитивните вистински титаносауруси, исто така, го задржале ноктот на предните нозе, но развиле екстремитети со широк мерач. Екстремитетите со широк мерач биле задржани од напредни титаносаури, патеки од кои се гледа широк мерач и недостаток на канџи на предната нога.[72] Повремено, се наоѓаат само патеки од предниот дел. Фалкингем и сор. [73] користеле компјутерско моделирање за да покаже дека ова може да се должи на својствата на подлогата. Овие треба да бидат исправни за да се зачуваат трагите.[74] Разликите во површината на задните екстремитети и предните екстремитети, а со тоа и притисокот на допирот со подлогата, понекогаш може да доведат до зачувување на патеките од предниот дел. Биомеханика и брзина Во студијата објавена во PLoS ONE на 30 октомври 2013 година, од Бил Селерс, Родолфо Корија, Ли Маргетс и др., Аргентиносаурус бил дигитално реконструиран за да го тестира своето движење за прв пат. Пред студијата, најчестиот начин за проценка на брзината бил преку проучување на коскена хистологија и ихнологија. Вообичаено, студии за хистологија на коските на сауропод и фокус на брзината на посткранијалниот скелет, кој има многу уникатни одлики, како што е зголемен процес на улна, широк лобус на илија, горната третина на бедрената коска навнатре и крајно јајцевидно феморално вратило. Овие одлики се корисни кога се обидувате да ги објасните моделите на патеките на гравипорталните животни. Кога студирате ихнологија за да ја пресметате брзината на сауропод, има неколку проблеми, како што е само обезбедување проценки за одредени постери поради пристрасност на зачувување, и подложени на многу повеќе проблеми со точност.[75] За да се процени одењето и брзината на Аргентиносаурусот, студијата извршила анализа на мускулно-скелетниот систем. Единствените претходни анализи на мускулно-скелетните органи биле спроведени на хоминоиди, терористички птици и други диносауруси. Пред да можат да ја направат анализата, тимот морал да создаде дигитален скелет на предметното животно, да покаже каде ќе има слоеви на мускулите, да ги пронајде мускулите и зглобовите и конечно да ги пронајде мускулните својства пред да го пронајде одењето и брзината. Резултатите од студијата за биомеханика откриле дека Аргентиносаурусот бил механички компетентен со максимална брзина од 2 м/с (5 на час) со оглед на големата тежина на животното и видот што неговите споеви биле способни да го носат.[76] Резултатите понатаму откриле дека многу поголеми копнени ’рбетници се можни, но ќе биде потребно значително реновирање на телото и можна доволна промена во однесувањето за да се спречи колапс на зглобот.[75] Големина на телото Сауроподите биле гигантски потомци на изненадувачки мали предци. Базалните, како што се Pseudolagosuchus и Marasuchus од Средниот тријас на Аргентина, тежеле приближно 1 килограм или помалку. Овие еволуирале во сауришија, која забележала брзо зголемување на големината на телото, иако попримитивните членови како Еораптор, Панфагија, Пантидрако, Сатурналија и Гуаибасаурус сè уште ја задржале умерената големина, веројатно под 10 килограми . Дури и со овие мали, примитивни форми, има значително зголемување на големината кај сауроподоморфите, иако скудни остатоци од овој период ја прават интерпретацијата претпоставка. Постои еден дефинитивен пример за мал изведен сауроподоморф: Анхисаурус, под 50 килограми , иако е поблиску до сауроподите од Платеосаурус и Риосасаурус, кои биле нагоре со тежина од 1 тон.[50] Еволуирајќи од сауроподоморфите (гуштероноголики), сауроподите биле огромни. Нивната џиновска големина веројатно резултирала од зголемена стапка на раст што е овозможено со тахиметаболична ендотермија, одлика што се развила во сауроподоморфи. Откако се разграниле во сауроподи, гуштероноголиките продолжиле стабилно да растат поголеми, со помали сауроподи, како диносаурусите од раната јура Барапасаурус и Котасаурус, еволуирајќи во уште поголеми форми како диносаурусите од средната јура Маменчисаурус и Патагосаурус . Одговарајќи на растот на сауроподи, нивните грабливци-тероподи растеле исто така, што е прикажано од целофизоидите како Алосаурусот од Германија .[50] Големина во НеосауроподаНеосауропода е сосема веројатно клад од диносауруси со најголеми димензии на телото што некогаш постоеле. Неколку исклучоци од помала големина се претпоставува дека се предизвикани од островското џуџестство, иако постои тренд во Титаносаурија кон помала големина. Титаносаурусите, сепак, биле едни од најголемите сауроподи досега. Освен титаносаурусите, клад на диплодоиди, група гиганти, наречени Дикреосауриди, се идентификувале со мала големина на телото. Дури и „џуџестите“ сауроподи се поголеми од 500 килограми, големина достигната само од околу 10% од сите видови цицачи.[50] Независен гигантизамИако сауроподите биле големи, огромната големина (40 тони или повеќе) била постигната независно во повеќе наврати од нивната еволуција. Многу гигантски форми постоеле во доцната јура (конкретно кимериџ), како што се турисаурусот Turiasaurus и диплодоцидите Марапунисаурус, Диплодокус и Баросаурус. Од почетокот до крајот на креда, гигантите како Сауропосејдон, Паралититан, Аргентиносаурус, Пуертасаурус, Antarctosaurus giganteus, Dreadnoughtus schrani, Нотоколосус и Футалогнкосаурус живееле, и сите тие се призати како титаносауруси. Еден ретко познат можен гигант е 'Хуангетитанскиот рујангенис, познат само од 3 метри долги ребра. Овие џиновски видови живееле од доцната јура до доцната Креда, се појавувале независно и живееле пред 85 милиони години.[50] Изоларно џуџе во сауроподиДва добро познати видови островски џуџиња на сауроподи се од периодот на креда Маџаросаурус (во еден момент бил оспорен неговиот идентитет како џуџе) и од периодот на јура, Европасаурус, и двата од Европа. И покрај тоа што овие сауроподи се мали, единствениот начин да се докаже дека се вистински џуџиња е преку студија за нивната коскена хистологија. Студија на Мартин Сандер и неговите колеги во 2006 година испитала единаесет единки од Europasaurus holgeri користејќи коскена хистологија и покажало дека малиот островски вид еволуирал преку намалување на стапката на раст на долгите коски во споредба со стапките на раст кај предците на копното.[77] Две други можни џуџиња се Рапетосаурус, кој постоел на островот Мадагаскар, изолиран остров во периодот креда, и Ампелосаурус, титаносаурус кој живеел на Пиринејскиот Полуостров во јужна Шпанија и Франција. Аманзија од Швајцарија исто така може да биде џуџе, но тоа допрва треба да се докаже.[50] Еден од најекстремните случаи на џуџест сауропод е Еуропасаурус, роднина на многу поголемите Камарасаурус и Брахиосаурус : тој бил само околу 6,2 метри долг.[78] Историја на откритиетоПрвите остатоци од фосили, сега признати како сауроподи, дошле од Англија и првично биле толкувани на различни начини. Нивната врска со другите диносауруси не била препознаена дури по нивното првично откритие.   Првиот фосил од сауропод кој бил научно опишан бил еден заб опишан како Rutellum implicatum.[79] Овој фосил бил опишан од Едвард Лујд во 1699 година, но не бил признат како џиновски праисториски влекач во тоа време.[80] Диносаурусите не би биле признати како група сè додека не поминало повеќе од еден век. Ричард Овен ги објавил првите современи научни описи на сауроподи во 1841 година, во книга и труд именувајќи ги Кардиодон и Цетиосаурус . Кардиодон бил познат само од два необични заби во форма на срце (од кои го добил и своето име), што не можело да се идентификува подалеку од фактот дека тие потекнуваат од претходно непознат голем влекач . Цетиосаурусот бил познат од малку подобри остатоци. Овен во тоа време помислил дека Цетиосаурусот е огромен морски влекач поврзан со современи крокодили, па оттука и неговото име, што значи „китски гуштер“. Една година подоцна, кога Овен го смислил името Диносаурија, тој не ги вклучил Цетиосаурусот и Кардиодонот во таа група.[81] Во 1850 година, Гедеон Мантел ја препознал диносауриската природа на неколку коски доделени од Овен на Цетиосаурусот. Мантел забележал дека коските на ногата содржат медуларна празнина, одлика на копнените животни. Овие примероци ги додели на новиот род Пелоросаурус и ги групирал заедно со диносаурусите. Сепак, Мантел сè уште не ја признавал врската со Цетиосаурусот.[46] Следниот пронајден сауропод што треба да се опише и погрешно да се идентификува како нешто друго освен диносаурус, бил збир на пршлени на колкот опишани од Хари Сили во 1870 година. Сили открил дека пршлените се многу лесно конструирани за нивната големина и содржат отвори за воздушни вреќи (пневматизација). Таквите воздушни вреќи биле во тоа време познати само кај птиците и птеросаурусите, а Сили сметал дека пршлените потекнуваат од птеросаурус. Новиот род го именувал Орнитопсис, или „птичјо лице“.[46] Кога Филипс ги опишал поцелосните примероци на Цетиосаурус во 1871 година, тој конечно го препознал животното како диносаурус поврзан со Пелоросаурус .[82] Сепак, дури во описот на нови, скоро целосни скелети на сауропод од Соединетите Држави (кои ги претставуваат Апатосаурусите и Камарасаурусите ) подоцна истата година се појавила целосна слика за сауроподи. Приближна реконструкција на комплетен скелет на сауропод била изготвена од уметникот Џон А. Рајдер, ангажиран од палеонтологот Коуп, врз основа на остатоците од Камарасаурус, иако многу одлики сепак биле неточни или нецелосни според подоцнежните откритија и биомеханичките студии.[83] Исто така, во 1877 година, Ричард Лидекер именувал друг роднина на Цетиосаурус, Титаносаурус, врз основа на изолиран пршлен.[46]  Во 1878 година, најпознатиот комплетен сауропод досега бил пронајден и опишан од Отниел Чарлс Марш, кој го нарекол Диплодокус. Со ова откритие, Марш исто така создал нова група која ги содржи Диплодокус, Цетиосаурус и нивниот сé поголем список на роднини за да ги разликува од другите поголеми групи на диносауруси. Марш ја нарекол оваа група Сауропода, или „стапала на гуштер“.[46] КласификацијаКласификацијата на сауроподите во голема мерка се стабилизира во последниве години, иако сè уште има некои несигурности, како што се поставувањето на Еухелоп, Хаплакантосаурус, Џобарија и Немегтосауриди. Следниот калдограм го направил Сандер и неговите колеги во 2011 година.[50]
Наводи
Литература
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||













Portal di Ensiklopedia Dunia













